Anna Katharina Franke  ,
Pasi Aatsinki,
Ville Hallikainen,
Esa Huhta,
Mikko Hyppönen,
Vesa Juntunen,
Kari Mikkola,
Seppo Neuvonen,
Pasi Rautio
,
Pasi Aatsinki,
Ville Hallikainen,
Esa Huhta,
Mikko Hyppönen,
Vesa Juntunen,
Kari Mikkola,
Seppo Neuvonen,
Pasi Rautio
Quantifying changes of the coniferous forest line in Finnish Lapland during 1983–2009
Franke A. K., Aatsinki P., Hallikainen V., Huhta E., Hyppönen M., Juntunen V., Mikkola K., Neuvonen S., Rautio P. (2015). Quantifying changes of the coniferous forest line in Finnish Lapland during 1983–2009. Silva Fennica vol. 49 no. 4 article id 1408. https://doi.org/10.14214/sf.1408
Highlights
- Volume of the growing stock of spruce and pine has increased in forests and in timber lines during the past 26 years
- Spruce stem numbers increased on average, while pine stem numbers remained stable and location-specific variation was observed
- Presuming that the ongoing trend of increasing temperature will remain, the enhanced forest regeneration and growth may result in extension of forests in the future.
Abstract
The boreal timber- and tree-line forests grow in harsh environmental conditions in their outermost distribution limit. Here even small environmental changes may cause dramatic changes in the distribution of tree species. We examined changes of the forest lines of Norway spruce (Picea abies (L.) H. Karst.) and Scots pine (Pinus sylvestris L.) in Finnish Lapland five times during 1983–2009. We monitored the number of stems and the volume of the growing stock in thirteen different locations in forest-line areas. The linear temporal trends and the variations of these response variables were used as indicators of a possible change during the study period. Spruce showed a significant increase both in the volume of the growing stock (up to 40% increase) and in the total stem number (up to 100% increase). A significant increase in the volume of the growing stock was observed in the pine data as well (up to 70% increase), whereas the stem number stagnated or even decreased. The results suggest that spruce needs favourable conditions to have an abundant regeneration, but after the establishment the seedlings seem to be more resistant against biotic and abiotic disturbances than pine seedlings. The increasing stand volume might result in a climate-related northward and upward extension of forests in the future. However, our results show that responses in the boreal forest line are species and location specific and a more favourable climate does not necessarily lead to an advance of the coniferous forest line.
Keywords
Pinus sylvestris;
Picea abies;
tree line;
forest regeneration;
environmental change;
stand volume
-
Franke,
Natural Resources Institute Finland (Luke), Management and Production of Renewable Resources, Eteläranta 55, FI-96301 Rovaniemi, Finland
E-mail
anna.franke@fau.de
- Aatsinki, Natural Resources Institute Finland (Luke), Management and Production of Renewable Resources, Eteläranta 55, FI-96301 Rovaniemi, Finland E-mail pasi.aatsinki@luke.fi
- Hallikainen, Natural Resources Institute Finland (Luke), Management and Production of Renewable Resources, Eteläranta 55, FI-96301 Rovaniemi, Finland E-mail ville.hallikainen@luke.fi
- Huhta, Natural Resources Institute Finland (Luke), Management and Production of Renewable Resources, Eteläranta 55, FI-96301 Rovaniemi, Finland E-mail esa.huhta@luke.fi
- Hyppönen, Natural Resources Institute Finland (Luke), Management and Production of Renewable Resources, Eteläranta 55, FI-96301 Rovaniemi, Finland E-mail mikko.hypponen@luke.fi
- Juntunen, The Sámi Education Institute, Menesjärventie 4, P.O. Box 50, FI-99871 Inari, Finland E-mail vesa.juntunen@sogsakk.fi
- Mikkola, Natural Resources Institute Finland (Luke), Economics and society, Eteläranta 55, FI-96301 Rovaniemi, Finland E-mail kari.mikkola@luke.fi
- Neuvonen, Natural Resources Institute Finland (Luke), Management and Production of Renewable Resources, P.O. Box 68, FI-80101, Joensuu, Finland E-mail seppo.neuvonen@luke.fi
- Rautio, Natural Resources Institute Finland (Luke), Bio-based business and industry, Eteläranta 55, FI-96301 Rovaniemi, Finland E-mail pasi.rautio@luke.fi
Received 16 June 2015 Accepted 15 July 2015 Published 18 August 2015
Views 164773
Available at https://doi.org/10.14214/sf.1408 | Download PDF

1 Introduction
The forest-line ecotone of the boreal zone, which separates treeless areas from closed boreal forests, is one of the Earth’s largest vegetation transition zones, extending more than 13 000 km around the Northern Hemisphere (Callaghan et al. 2002). Boreal vegetation has adapted to adverse environmental conditions, such as a short growing season, low temperatures and nutrient supply as well as sparse light during winter (Kallio et al. 1986; Heikkinen et al. 2002). In Finnish Lapland, for example, the growing season lasts less than four months during which trees need to pass through all vegetative (bud break, annual growth) and reproductive (flowering, cone production) stages (Seo et al. 2010). In northern boreal forests, the temperature during the growing season is the main factor that influences the growth and survival of coniferous species (Esteban and Jackson 2000; Heikkinen et al. 2002; Juntunen et al. 2002). Even small temperature changes might cause strong response e.g. in growth or bud burst (Grace et al. 2002; Linkosalo et al. 2009; Høgda et al. 2013; Salminen and Jalkanen 2015). Favourable conditions also allow abundant seed production leading to higher regeneration and seedling establishment (Hilli et al. 2008). Consequently, tree survival and growth in forest-line areas can be used as indicators for a change of the environmental conditions (Karlsen et al. 2005).
The analysis of the sub-fossil Scots pine (Pinus sylvestris L.) trunks coming from north of the current coniferous forest line in Finland suggests a warmer-than-present climate between 8 000 and 4 000 BP, and a forest line located at higher altitudes and latitudes compared to the present (Seppä et al. 2002; Kultti et al. 2006). However, the pine forests retreated during a subsequent colder period to the present lower altitudes and more southern latitudes. Hence a warming climate might result in an advance of the current forest line to approach the previous distribution limit (Aakala et al. 2014). This makes forest-line monitoring important because recently rising temperatures and prolonged growing-seasons as well as varying amounts of precipitation have been observed in Finland (Klein Tank et al. 2002; Tietäväinen et al. 2010; Mikkonen et al. 2014).
As the latest decade (2001–2010) was the warmest one in Finland since the beginning of temperature measurements (starting in the 1840’s, Finnish Meteorological Institute 2014; Mikkonen et al. 2014; see also Fig. 1), we expect to see an advance of the forest line assuming that the regeneration correlates positively with temperature. During the last 50 years, the climate has been favourable enough for the regeneration of forests in the forest-line areas (Holtmeier 2005; Hyppönen et al. 2013). Previous studies in our study area have indicated that seedling establishment has been at least satisfactory and the mortality of seedlings has remained rather low (Juntunen et al. 2002; Juntunen and Neuvonen 2006).
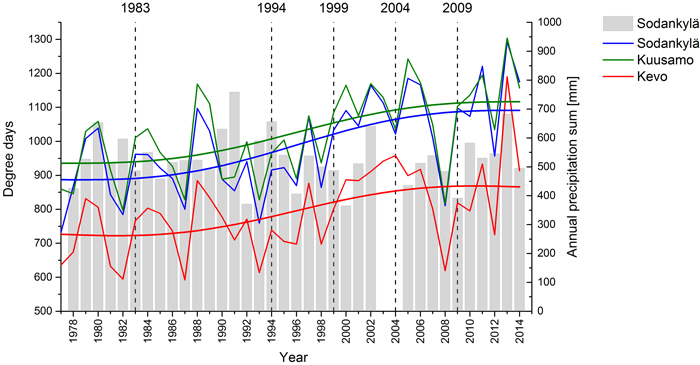
Fig. 1. Effective temperature sums (+4 °C threshold) in Kuusamo, Sodankylä and Kevo and annual precipitation sums (columns) in Sodankylä from 1977 to 2013. The climate datasets (thin lines, columns) are provided in a yearly resolution by European Climate Assessment & Dataset (Klein Tank et al. 2002) and smoothed by using a Fast Fourier Transformation (thick lines).
Several environmental factors determine the regeneration and death of trees at the forest line affecting the balance between increase and decrease of the volume of the growing stock in a stand. These factors are assumed to be related to prevailing climatic conditions at the forest line. Favourable climatic conditions support seedling production and tree growth. However, a changing climate may also create adverse conditions, which may increase death of trees via disturbances, for example, by increasing storm and snow damage (Gregow et al. 2011), large temperature variations (Heikkinen et al. 2002) and providing favourable conditions for the outbreak of pests and diseases (Jalkanen 2003; Holtmeier 2005).
In this paper we investigate changes at forest lines of Scots pine and Norway spruce (Picea abies (L.) H. Karst.) during a 26-year period. We monitored the number of stems and the volume of the growing stock in forest-line areas in Finnish Lapland during 1983–2009. The linear temporal trends and the variations of these response variables were used as indicators of a possible change. First responses of the coniferous forest lines to a changing environment have been already observed in the first part of the study during 1983–1999 (Juntunen et al. 2002). Here we predict an increase in the number of stems and the volume of the growing stock of pine and spruce in the forest-line ecotone in response to the increased summertime temperatures during the past decade.
2 Materials and methods
2.1 Study area, sampling design and measurements
In our study area in Finnish Lapland, coniferous forest lines are alpine or polar-alpine and they occur over an area spanning 400 km in the south-north direction (Juntunen et al. 2002). These forest lines vary in nature from the Southern to the Northern Lapland. In Southern Lapland forest lines are clearly alpine forest lines and occur on scattered fells in the midst of continuous boreal forest. The northern forest lines are found on higher altitudes as well but are mostly a result of harsh growing conditions on high latitudes, where the boreal forest meets the mountain birch forest zone (Betula pubescens Ehrh.) forming the outermost limit of forest vegetation (Hämet-Ahti 1963). The regional climate is mostly (sub)continental with cold winters and relatively warm summers (Tuhkanen 1980).
The forest-line monitoring project is being carried out by the Finnish Forest Research Institute (from beginning of 2015, Natural Resources Institute Finland) and the universities of Helsinki, Oulu and Turku (Kallio et al. 1986). To monitor the forest lines in Finnish Lapland the area was divided into four regions on the basis of predominant tree species and geographical areas (Juntunen et al. 2002). The pine-dominated areas were divided into a northern and a southern region (P-N and P-S), and the spruce-dominated areas into a western and an eastern region (S-W and S-E). Each region includes two to four locations, giving a total of 13 locations (Scots pine 8 and Norway spruce 5) in northern Finland (Fig. 2). The 13 locations were carefully chosen to cover the whole area of Lapland but at the same time avoiding locations where the topography or soil factors would affect the results. At the same time places where marks of past disturbances were present were excluded. The mean age of the studied stands was not possible to record as the age of seedlings, saplings and trees varied from 1-year old seedlings to several hundred years old trees. Hence we presume that the age of the studied stands in the beginning of the monitoring was the same as the age of the dominant trees. According to the sample cores taken from the old trees for another project in the studied locations the stand age varied from 200 to 400 years in forest and timber-line zones and from 150 to 200 years in tree-line zone (Fig.3).
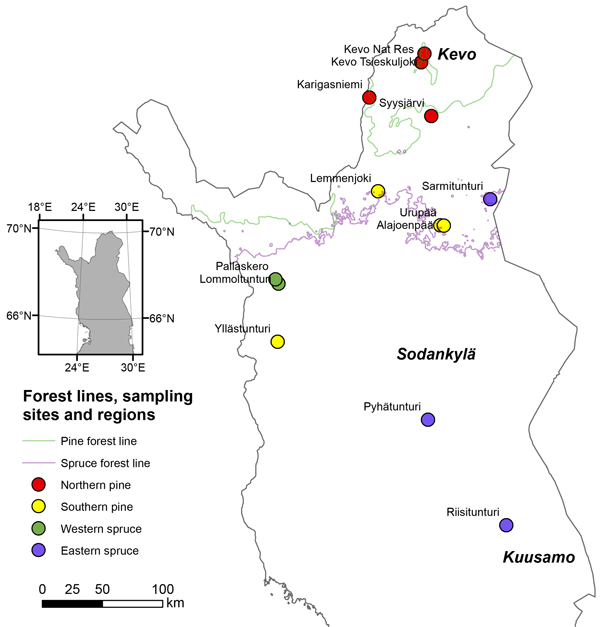
Fig. 2. Monitoring locations of spruce and pine in Finnish Lapland as well as the forest lines of both species. The locations were divided on the basis of predominant species and geographical areas into a northern and southern region of pine and into a western and eastern region of spruce.

Fig. 3. Examples of the forest stands in the a) forest zone (Yllästunturi), b) timber-line zone (Kevo) and c) tree-line zone (Urupää) (Photos by Seppo Neuvonen and Anna Franke).
Each of the locations consists of a system of three rows of three circular sample plots along an altitudinal gradient from forest to tree line (Fig. 4). These three rows were located in at least 100 meters distance from each other and differed in vegetation pattern, tree height and stand density and can therefore be considered as independent stands (Fig. 5, Table 1). The number of analyzed forest stands is therefore 39. The distance between the centres of adjacent monitoring plots was 40 m. The row in the highest altitude was established within the tree-line zone, just below the tree line, where the distance between solitary trees was higher than 2 m but did not exceed 100 m. The second row was established in timber-line zone, which is defined here as the altitudinal limit at which the forest canopy closure ceases (Hustich 1948). The lowest row was then established in a forest stand representing the characteristics of the forest zone. It was placed so that its vertical distance from the timber-line row was at least 20 m, or its horizontal distance over 100 m, or both. The forest-line area in this study contains the entire transition belt in general, extending from closed coniferous forests up to fell heaths. The total number of the plots in the experiment is 117 (nine plots in each 13 locations). Most of the plots have an area of 300 m2 (radius 9.8 m); however, in the places where there were only a few trees, 500 m2 plots (radius 12.6 m) were used (Table 1). The average of three plots in each zone was used in the statistical analysis, because it gives a reliable estimate for forest stand level demographic processes in our study locations. Therefore, the original values (counts and measurements) per sample plot have been transformed to the values per hectare.
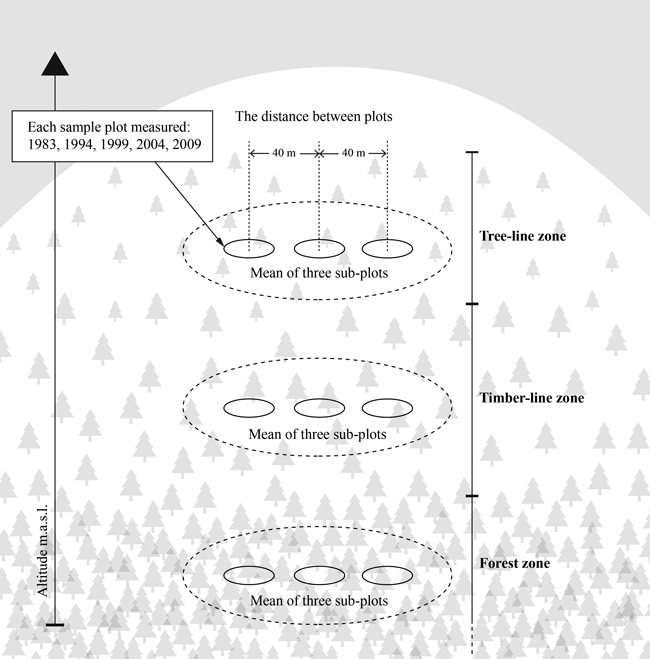
Fig. 4. Sampling design in each of the 13 locations. The sampling was done on three sample plots in forest and timber- and tree-line zones in 1983, 1994, 1999, 2004 and 2009.

Fig. 5. Example of a sampling location (Sarmitunturi) showing the position of the timber-line and tree-line zones (Photo by Raimo Sutinen). The forest zone is located further downhill and therefore not shown in the photo.
| Table 1. Coordinates, altitude, temperature sum, and horizontal distance between the zones in each location. Coordinates and temperature sums are focused on the timber-line zones. The average temperature sums (+5 °C threshold) were provided in a climatic grid (1961–90 standard period) by using the model of Ojansuu and Henttonen (1983). | |||||||||
| Altitude (m a.s.l.) | Horizontal distance (m) between | ||||||||
| Location | Latitude | Longitude | Dominant conifer1 | Forest zone | Timber line | Tree line | Temperature sum (d.d. 5 °C) | Forest and timber line | Timber line and tree line |
| Kevo Tsieskuljoki | 69°41’ N | 26°59’ E | P | 110 | 180 | 190 | 773 | 310 | 180 |
| Kevo, nat. res | 69°45’ N | 27°03’ E | P | 185 | 210 | 225 | 741 | 230 | 100 |
| Karigasniemi | 69°25’ N | 25°53’ E | P | 225 | 275 | 420 | 770 | 840 | 810 |
| Syysjärvi | 69°17’ N | 27°12’ E | P | 215 | 220 | 225 | 781 | 1280 | 11460 |
| Lemmenjoki | 68°43’ N | 26°06’ E | P | 285 | 375 | 415* | 782 | 460 | 130 |
| Urupää | 68°28’ N | 27°26’ E | P | 315 | 340 | 370* | 745 | 140 | 210 |
| Alajoenpää | 68°28’ N | 27°22’ E | P | 340 | 380 | 400* | 748 | 250 | 270 |
| Ylläs | 67°34’ N | 24°11’ E | P | 380 | 410 | 465* | 900 | 240 | 210 |
| Lommoltunturi | 68°00’ N | 24°09’ E | S | 410 | 445 | 465* | 874 | 250 | 120 |
| Pallaskero | 68°02’ N | 24°06’ E | S | 465 | 480 | 500* | 892 | 220 | 110 |
| Sarmitunturi | 68°39’ N | 28°23’ E | S | 340 | 370 | 410* | 842 | 150 | 160 |
| Pyhätunturi | 67°01’ N | 27°07’ E | S | 380 | 400 | 420 | 914 | 300 | 260 |
| Riisitunturi | 66°13’ N | 28°33’ E | S | 420 | 440 | 460* | 943 | 120 | 190 |
| 1 Dominant conifer species: P = pine, S = spruce. * Indicates that a larger sample plot area (500 m2) has been used. | |||||||||
After the establishment of the plots in 1983, monitoring was repeated in 1994, 1999, 2004 and 2009. The monitoring on the plots includes the number of living trees ( ≥ 2 m height), saplings ( < 2 m and ≥ 1.3 m) and seedlings (height < 1.3 m) as well as measurements of the height and diameter at breast height of all trees within a plot to compute the volume of the growing stock with the KPL-software (Heinonen 1994). The volume of the growing stock includes only trees over 2 meters, (i.e. not seedlings and saplings). In this study an increasing growing stock denotes that the volume increment of a forest stand exceeds the loss of volume due to mortality and represents, therefore, a positive in-balance of growth.
The climate datasets are provided in a yearly resolution by European Climate Assessment & Dataset from 1977 to 2014 for Kevo, Kuusamo and Sodankylä (Klein Tank et al. 2002, http://www.ecad.eu). To characterize the climatic conditions during the growing season we used the effective temperature sum expressed in degree days and the annual precipitation sum. The degree day as a linear temperature sum is based on daily mean temperature minus the threshold value of +4 °C, used by the database for continental Europe. The results were not fitted to the threshold value for Finland (+5 °C) in this context because the threshold does not affect the detected trends and it is likely to cause only minor differences in the resulting temperature sums. Since there were no meteorological stations near the study plots, we used a climatic grid of temperature sums (+5 °C threshold) to receive the temperature sum for each location. The grid data was computed by using the model of Ojansuu and Henttonen (1983) for averaging the temperature sums within the period from 1961–90.
2.2 Statistical analysis
Linear mixed models were used in the modelling of the data. Separate models were calculated for two response variables: 1) The volume of the growing stock (m3 ha–1) and 2) total number of trees, saplings and seedlings (ha–1), in the following used as “stem number”. These two models were calculated separately for Scots pine- and Norway spruce -dominated locations. The response variables were calculated by summing the values (number of stem and the volume of the growing stock) of spruces and pines in each sample plot. Logarithmic (ln) transformation was used for the stem number and square-root transformation for the volume of the growing stock. These transformations yielded the most unbiased residuals (no trends, consistent variances) for the models. The model for the volume of the growing stock can be expressed as following:
, where
sqrt(ŷijt) = Estimated square root of the volume of the growing stock (separate models for the locations dominated by Scots pine and Norway spruce)
x1(i) = Fixed predictor of region (levels: northern, southern (Scots pine), western, eastern (Norway spruce))
x2(ij) = Fixed predictor of zone (levels: forest, timber line, tree line)
x3(ijt) = Fixed predictor of time (0 – 26 years, 0denotes year 1983 and 26 year 2009 respectively)
β0,..., β5 = Coefficients of the fixed effects and interactions of the fixed effects
μ1(i) = Random effect of location (Scots pine: 8 locations; Norway spruce: 5 locations)
μ2(ij) = Random effect of zone nested within location
ɛijt = Random error (t, denoting repeated measures), nested within zone (j). Autoregressive (AR-1) error, heterogeneous variances for each repeated measures time points were estimated.
Compound symmetry was used as the random error structure in the log-transformed models for the stem number, instead of the autoregressive structure based on likelihood-ratio tests and residuals (minimizing bias). The same model structure based on the experimental design was used in all of the models and all the coefficients and their significances were reported, including the two-way interaction terms (formula 1). The three way interaction of region, zone and time could not be tested due to convergence problems.
In addition we tested if the coefficients of time by region and by zone differ from 0(results are presented in Table 4, null hypothesis: β = 0). These tests indicate if the stem number as well as the volume of the growing stock has increased, decreased or remained stable during the 26-year period. The tests and the coefficients in the tables 2–4 were generated by the models using the transformed response. However, the fitted values presented in the text and figures were transformed back to the original scale (exponential and power transformations). The values of the exponential transformations were corrected using Snowdon’s (1991) ratio estimator and those of power transformation by adding variances of the random parts to the transformed values.
| Table 2. The estimates and Likelihood-ratio tests of the models for Norway spruce, t-values for the coefficients and χ2 – values for the Likelihood ratio tests are presented in t/Chisq. Tests show the significance of the variables in the model and the significance of the coefficients related to the reference categories (given in parentheses). A non-significant interaction term means that the coefficients do not differ from the coefficient of reference category. | ||||||
| Variable / term | Coefficient | Std. error | Df | t/Chisq | p | |
| Norway spruce, number of trees and seedlings ha–1 | ||||||
| Intercept | 7.000 | 0.376 | 56/1 | 18.62/346.54 | 0.000 | |
| Region (ref. Western) | 1 | 4.28 | 0.039 | |||
| -Eastern | -0.854 | 0.412 | 3 | -2.07 | 0.130 | |
| Zone (ref. Forest) | 2 | 3.70 | 0.157 | |||
| -Timber line | 0.333 | 0.342 | 8 | 0.97 | 0.359 | |
| -Tree line | -0.325 | 0.342 | 8 | -0.95 | 0.370 | |
| Time | 0.018 | 0.010 | 56/1 | 1.77/3.13 | 0.077 | |
| Region*Time | 1 | 1.81 | 0.179 | |||
| -Eastern*Time | -0.013 | 0.010 | 56 | -1.35 | 0.184 | |
| Zone*Time | 2 | 3.49 | 0.175 | |||
| -Timber line*Time | 0.006 | 0.012 | 56 | 0.48 | 0.630 | |
| -Tree line*Time | 0.021 | 0.012 | 56 | 1.80 | 0.077 | |
| Random effects | Estimate | Cl 95% | ||||
| Location | 0.107 | 0.007 – 1.682 | ||||
| Zone nested Location | 0.187 | 0.061 – 0.579 | ||||
| Error (compound symmetry) | 0.141 | 0.098 – 0.205 | ||||
| Norway spruce, volume of tree stock, m3ha–1 | ||||||
| Intercept | 7.970 | 1.073 | 56/1 | 7.43/55.19 | 0.000 | |
| Region (ref. Western) | 1 | 0.65 | 0.420 | |||
| -Eastern | -1.031 | 1.278 | 3 | -0.81 | 0.479 | |
| Zone (ref. Forest) | 2 | 67.50 | 0.000 | |||
| -Timber line | -2.056 | 0.717 | 8 | -2.87 | 0.021 | |
| -Tree line | -5.812 | 0.717 | 8 | -8.10 | 0.000 | |
| Time | 0.043 | 0.010 | 56/1 | 4.27/18.21 | 0.000 | |
| Region*Time | 1 | 2.85 | 0.091 | |||
| -Eastern*Time | -0.017 | 0.010 | 56 | -1.69 | 0.097 | |
| Zone*Time | 2 | 8.10 | 0.017 | |||
| -Timber line*Time | 0.012 | 0.012 | 56 | 1.04 | 0.301 | |
| -Tree line*Time | -0.021 | 0.012 | 56 | -1.77 | 0.082 | |
| Random effects | Estimate | Cl 95% | ||||
| Location | 1.530 | 0.197 – 11.853 | ||||
| Zone nested within Location | 1.186 | 0.443 – 3.177 | ||||
| Error (ARH-1, phi = 0.55) | 0.398 | 0.165 – 0.957 | ||||
| Table 3. The estimates and Likelihood-ratio tests of the models for Scots pine, t-values for the coefficients and χ2 – values for the Likelihood ratio tests are presented in t/Chisq. Tests show the significance of the variables in the model and the significance of the coefficients related to the reference categories (given in parentheses). A non-significant interaction term means that the coefficients do not differ from the coefficient of reference category. | ||||||
| Variable / term | Coefficient | Std. error | Df | t/Chisq | p | |
| Scots pine, number of trees and seedlings ha–1 | ||||||
| Intercept | 6.577 | 0.695 | 90/1 | 9.43/88.90 | 0.000 | |
| Region (ref. Northern) | 1 | 0.84 | 0.359 | |||
| -Southern | 0.729 | 0.795 | 6 | 0.92 | 0.395 | |
| Zone (ref. Forest) | 2 | 10.12 | 0.006 | |||
| -Timber line | -1.040 | 0.710 | 14 | -1.47 | 0.165 | |
| -Tree line | -2.256 | 0.710 | 14 | -3.18 | 0.007 | |
| Time | 0.012 | 0.009 | 90/1 | 1.36/1.85 | 0.174 | |
| Region*Time | 1 | 12.95 | 0.000 | |||
| -Southern*Time | -0.032 | 0.009 | 90 | -3.60 | 0.001 | |
| Zone*Time | 2 | 0.91 | 0.635 | |||
| -Timber line*Time | -0.010 | 0.011 | 90 | -0.89 | 0.377 | |
| -Tree line*Time | -0.008 | 0.011 | 90 | -0.74 | 0.464 | |
| Random effects | Estimate | Cl 95% | ||||
| Location | 0.591 | 0.050 – 7.018 | ||||
| Zone nested Location | 1.876 | 0.882 – 3.990 | ||||
| Error (compound symmetry) | 0.185 | 0.138 – 0.248 | ||||
| Scots pine, volume of tree stock, m3ha–1 | ||||||
| Intercept | 7.124 | 1.166 | 90/1 | 6.11/37.34 | 0.000 | |
| Region (ref. Northern) | 1 | 0.00 | 0.997 | |||
| -Southern | -0.005 | 1.296 | 6 | -0.00 | 0.997 | |
| Zone (ref. Forest) | 2 | 24.57 | 0.000 | |||
| -Timber line | -3.665 | 1.247 | 14 | -2.94 | 0.011 | |
| -Tree line | -6.145 | 1.247 | 14 | -4.93 | 0.000 | |
| Time | 0.054 | 0.015 | 90/1 | 3.64/13.27 | 0.000 | |
| Region*Time | 1 | 0.50 | 0.478 | |||
| -Southern*Time | -0.011 | 0.015 | 90 | -0.71 | 0.479 | |
| Zone*Time | 2 | 5.47 | 0.065 | |||
| -Timber line*Time | 0.026 | 0.018 | 90 | 1.43 | 0.155 | |
| -Tree line*Time | -0.016 | 0.018 | 90 | -0.89 | 0.377 | |
| Random effects | Estimate | Cl 95% | ||||
| Location | 1.287 | 0.084 – 19.753 | ||||
| Zone nested within Location | 4.968 | 2.348 – 10.513 | ||||
| Error (ARH-1, phi = 0.91) | 1.325 | 0.533 – 3.294 | ||||
| Table 4. Significances (p-values) of asymptotical tests for the coefficients of the interaction terms (Time*Region or Time*Zone) in the models. Hypothesis is that the coefficients = 0. The arrows denote either a significant increasing or decreasing change in time, or non-significant change (flat arrow). | ||||
| Coefficient | Scots pine, number of trees ha–1 | Scots pine, volume of tree stock, m3ha–1 | Norway spruce, number of trees ha–1 | Norway spruce, volume of tree stock, m3ha–1 |
| Region: northern (pine), western (spruce) | 0.327 → | 0.000 ↗ | 0.000 ↗ | 0.000 ↗ |
| Region: southern (pine), eastern (spruce) | 0.000 ↘ | 0.000 ↗ | 0.026 ↗ | 0.000 ↗ |
| Zone: forest | 0.590 → | 0.000 ↗ | 0.127 → | 0.000 ↗ |
| Zone: timber line | 0.078 → | 0.000 ↗ | 0.041 ↗ | 0.000 ↗ |
| Zone: tree line | 0.120 → | 0.011 ↗ | 0.000 ↗ | 0.086 → |
In addition to the linear trends, the second-order polynomial of time and its interactions with region and zone were tested as the additional terms in the models for the volume of growing stock, but they were not significant at 5% risk level. Thus, only the linear trends are presented here. Significant polynomial terms would indicate an accelerative increase or decrease in the volume of growing stock.
R statistical environment (R Core Team 2013) was used in the modelling. Used packages were nlme (Pinheiro et al. 2013), Car (Fox and Weisberg 2011) and lsmeans (Lenth 2014).
3 Results
We detected a significant increase in the stem number of spruce in timber- and tree-line zones, whereas the forest zone remained stable (Fig. 6a, Tables 2 and 4). Even though the stem number of spruce increased in both western and eastern regions the increase was much higher in the west (on average 1 389 stems ha–1 in 1983 and 2 826 ha–1 in 2009, i.e. over 100% increase) compared to the east (591 stems ha–1 in 1983 and 850 ha–1 in 2009, i.e. 44% increase). The volume of the spruce growing stock (m3 ha–1) showed also increasing trends over the whole observation period in all zones, although the increase in the tree-line zone was statistically only marginally significant (Fig. 6b, Tables 2 and 4). When comparing the western and the eastern region, the western part indicated a higher gain of volume of 40% (from 31.5 m3 ha–1 in 1983 to 44.0 m3 ha–1 in 2009) compared to the 27% in the eastern part (from 21.6 m3 ha–1 to 27.4 m3 ha–1).
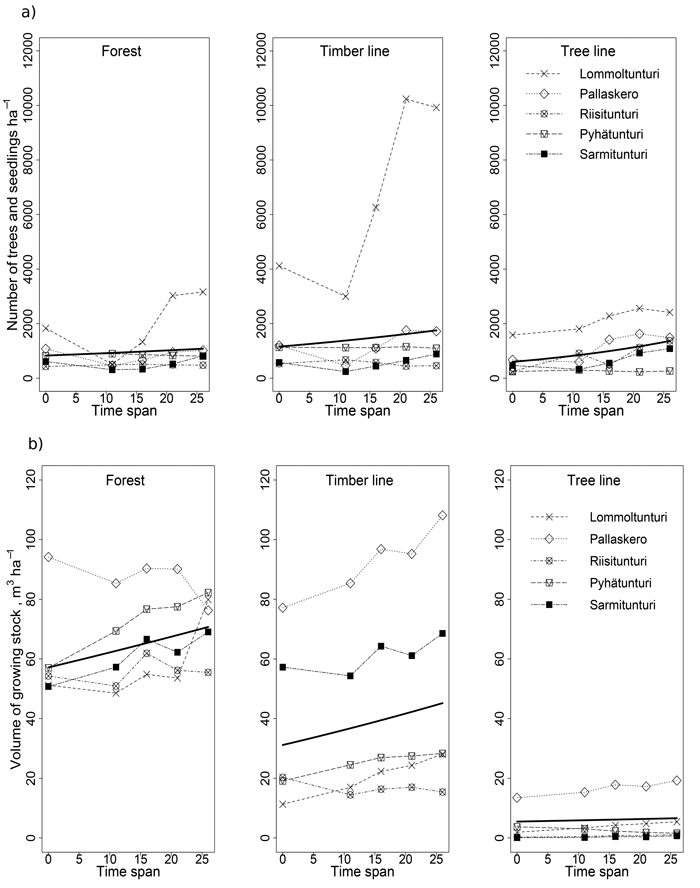
Fig. 6. The stem number ha–1 (a) and volume of growing stock m3 ha–1 (b) of spruce during the study in each location (raw data) as well as the predicted values of the models for each zone (thick line).
On the contrary to spruce, the stem number of pine showed no clear trends in any of the zones when all the locations are considered (Fig. 7a, Tables 3 and 4). When trends for the northern and southern pine regions were studied separately, a statistically significant decline in the pine number of 49% from 816 stems ha–1 in 1983 to 414 stems ha–1 in 2009 was found in the southern regions. The observed low increase of 17% from 394 stems ha–1 to 462 stems ha–1 in the northern region is not significant.
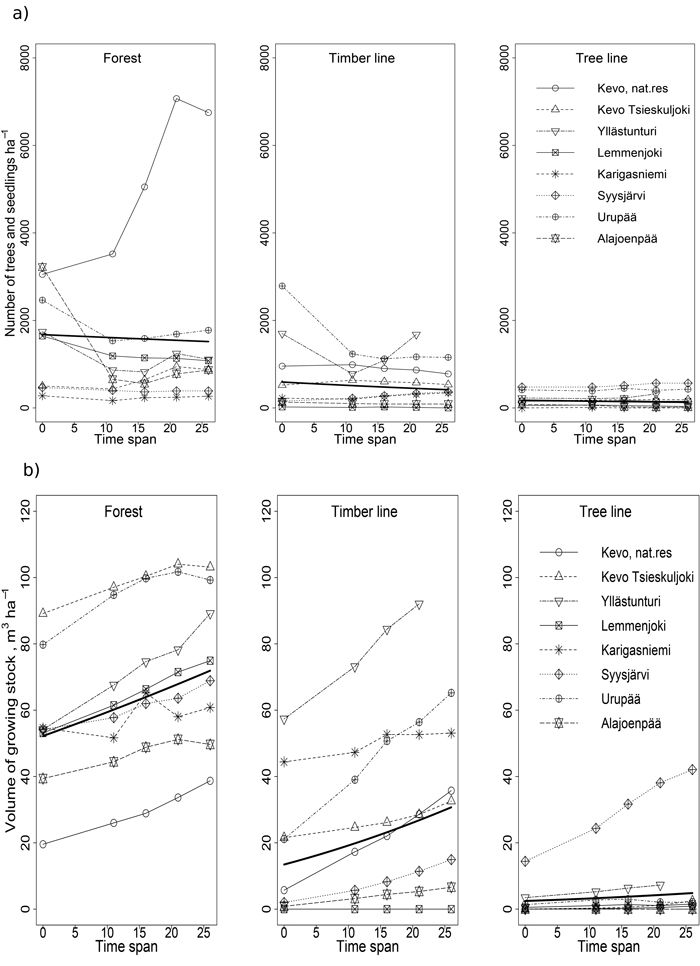
Fig. 7. The stem number ha–1 (a) and volume of growing stock m3 ha–1 (b) of pine during the study in each location (raw data) as well as the predicted values of the models for each zone (thick line).
Even though the number of the pines showed no clear overall change, the volume of the pine growing stock (m3 ha–1) increased significantly in all zones in the northern and the southern regions (Fig. 7b, Tables 3 and 4). Northern and southern distribution areas revealed an increased volume of 70% (from 18.6 m3 ha–1 to 33.4 m3 ha–1) in the north and 56% (from 19.5 m3 ha–1 to 30.4 m3 ha–1) in the south during the study period. A noteworthy fact is that even though there was a lot of variation between the locations the trend was, especially in the forest zone and in the timber line, positive in all locations.
4 Discussion
Our results indicated an increase in the volume of the growing stock of both spruce and pine in every zone and region, except in the case of spruce in the tree-line zone. An increasing growing stock here means that changing environmental conditions support higher reproduction, increment and survival of trees compared to the rate of dieback, leading to an advancing forest line. In other words, the studied stands were not in their climax balance, in which reproduction and growth are levelled out by death of trees, but a positive imbalance exists. The fact that the volume of the growing stocks was, on average, increasing gives a reason to assume that environmental conditions affecting growing stocks have, in general, improved since the first inventory in 1983. Temperature sums in Kuusamo, Sodankylä and Kevo as well as the amount of precipitation have risen during the study period in comparison to their long term annual mean values (1977–2013), especially during the period of 1994 to 2013 (Fig. 1). On the other hand, there was no clear indication for acceleration of the growing stock during the past decade that was the warmest decade in Finland ever recorded. In fact, in some of our study locations the growing stock has diminished during the last decade. This suggests that in addition to summer time temperature other factors have also had a strong impact on the growing stock. Kauppi et al. (2014) studied the effect of climate warming on boreal forests since 1960 and found out that in Finnish Lapland, where the annual growing degree days (GDD) have increased over 20% since 1960, the proportion of forest growth attributable to warming was around 43% whereas the rest was unrelated to warming. Kauppi et al. (2014) explained the part unrelated to warming by the factors such as forest management, increased nitrogen deposition and CO2 concentration when the whole Finland is considered. In our study locations, however, forests are not managed and the nitrogen deposition is very low (Mustajärvi et al. 2008). All this suggests that the trends seen in the growing stocks in our study locations are a response to interactions of warmer and longer growing season as well as increased CO2 with other abiotic and biotic factors.
On the contrary to the growing stock, the number of stems, including trees, saplings and seedlings, revealed a species and location specific response to changing environmental conditions. The significant increase in the number of spruce stems in the tree- and timber-line zones suggests that the end of the 20th century obviously provided favourable conditions for good seed years followed by good seedling establishment. The modest gain of the stem number in the forest zone is most likely a result of a between-tree competition for nutrients and light as well as of thick moss and humus layers which hinder regeneration (Juntunen and Neuvonen 2006).
Once spruce seedlings and saplings have been established successfully they seem to be more tolerant to disturbances. With regard to a significant increase of stem numbers, growth conditions near tree and timber lines seem to be favourable enough to enhance seeding, seedling establishment and survival of trees of all age classes. Supposing that prolonged growing seasons as well as higher temperatures in summer and winter times are at least partly responsible for the observed development, a northward and upward transition of spruce forest line can be expected where the geological factors, such as nutrient supply, allow this (Hyppönen et al. 2003; Sutinen et al. 2012).
In contrast to spruce, there was not a clear trend in the number of pine stems. When considering geographical regions separately, the stem number of pine in the southern part of the study area (locations Urupää, Alajoenpää and Yllästunturi) decreased especially during the first monitoring interval (1983–1994), but it is obvious that generally no large changes have taken place during the last 15 years (Fig. 7a). The strong reduction in the stem numbers in southern locations was mainly due to the high mortality of pine seedlings during 1983–2004 (Juntunen et al. 2002; Juntunen and Neuvonen 2006). Pine diebacks in the southern locations are a consequence of multiple biotic factors. Pines in these locations are often hit by the fungal diseases Scleroderris canker (Gremmeniella abietina (Lagerb.) Morelet) and the snow blight (Phacidium infestans P. Karst), as well as insects, such as the European pine sawfly (Neodiprion sertifer Geoffr.) (Niemelä et al. 1987; Juntunen and Neuvonen 2006; Holtmeier and Broll 2011). The European pine sawfly is predicted to have more outbreaks also in Finnish Lapland due to a reduced mortality of the eggs in milder winter temperatures (Virtanen et al. 1996; Neuvonen et al. 1999; Veteli et al. 2005).
Another biotic factor causing damage to pine seedlings is trampling by reindeer (Rangifer tarandus tarandus L.), when it is grazing. The reindeer population of Finland has more than doubled from about 100 000 reindeer in the beginning of the 1900s up to 265 000 in early 1990 (Väre et al. 1996) after which it has decreased to 197 000 in 2010 (Mattila 2014 a, b). Reindeer graze the reindeer lichens (Cladonia spp.) in pine dominated forests in winter that causes additional disturbance particularly to pine seedlings (Heikkinen et al. 2002; Holtmeier 2005; Vajda and Venäläinen 2005; Holtmeier and Broll 2011). Spruce grows in forests where lichens have only a minor role in understory and is therefore only marginally influenced by reindeer grazing.
Besides above biotic factors, seedlings are threatened by mild winters with an early start of the growing season and large temperature variations during late autumn and early spring, which influence their frost-hardiness (Cannell and Smith 1986). The risk of frost-damage induced mortality of seedlings is predicted to increase due to a warming climate and milder winter temperatures (Hänninen 1991; Leinonen 1995; Repo et al. 1996). Once trees have passed the seedling and sapling stage they have to survive from the stress of snow loading. Extreme crown-snow loading is the major reason for tree breakage at timber line in northern Finland (Marchand 1987), particularly at high altitudes and in old age-classes of the trees (Jalkanen and Konôpka 1998; Lehtonen et al. 2014). Spruce is, due to its slim crown shape, better protected against massive snow loading, whereas the shape of pine crown facilitates snow damage.
To sum up, our results show that the forests in the timber-line ecotone in Finnish Lapland are increasing in the volume of growing stock and tree number most likely due to favourable climatic conditions during the past decades. Presuming that the ongoing trend of increasing temperature will remain, the enhanced forest regeneration and growing stock in the timber- and tree line may result in a northward and upward extension of forests in the future. However, this process seems to be species specific, and in places where the pine is the main species, biotic factors or snow damage might overrule the effects of otherwise favourable environmental conditions. To evaluate the effect of abiotic and biotic factors more precisely, more detailed studies on cause-effect relationships are needed. However, results of the present monitoring study provide a valuable tool for decision-makers in forest policy, as well as a basis for further studies on factors influencing forest-line dynamics. Hence, the continuation of similar long-term studies is important to enable a reliable estimation of environmental changes in the forest-line ecotone.
Acknowledgements
This study would not have been possible without the great efforts that field assistants working in field stations of Finnish Forest Research Institute (Metla) as well as universities of Oulu, Turku and Helsinki have provided during the 26 years of monitoring. Risto Jalkanen and a peer in Peerage of Science gave valuable comments that helped to improve the manuscript. Jouni Hyvärinen designed the Figure 4. Raimo Sutinen kindly provided the photo for Figure 5.
References
Aakala T., Hari P., Dengel S., Newberry S.L., Mizunuma T., Grace J. (2014). A prominent stepwise advance of the tree line in north-east Finland. Journal of Ecology 2014: 1–10. http://dx.doi.org/ 10.1111/1365-2745.12308.
Callaghan T.V., Crawford R.M., Eronen M., Hofgaard A., Payette S., Rees W.G., Skre O., Sveinbjörnsson B., Vlassova T.K., Werkman B.R. (2002). The dynamics of the tundra-taiga boundary: an overview and suggested coordinated and integrated approach to research. AMBIO, Special Report 12: 3–5.
Cannell M.G.R., Smith R.I. (1986). Climatic warming, spring budburst and frost damage on trees. Journal of Applied Ecology 23: 177–191.
Esteban G.J., Jackson R.B. (2000). Global controls of forest line elevation in the northern and southern hemispheres. Global Ecology and Biogeography 9: 253–268. http://dx.doi.org/ 10.1046/j.1365-2699.2000.00162.x.
Finnish Meteorological Institute (2014). http://en.ilmatieteenlaitos.fi/. [Cited 14 Nov 2014].
Fox J., Weisberg S. (2011). An {R} companion to applied regression, second edition. Sage Publications, Thousand Oaks, CA. http://socserv.socsci.mcmaster.ca/jfox/Books/Companion. [Cited 15 Oct 2011].
Grace J., Berninger F., Nagy L. (2002). Impacts of climate change on the tree line. Annals of Botany 90: 537–544. http://dx.doi.org/10.1093/aob/mcf222.
Gregow H., Peltola H., Laapas M., Saku S., Venäläinen A. (2011). Combined occurrence of wind, snow loading and soil frost with implications for risks to forestry in Finland under the current and changing climatic conditions. Silva Fennica 45(1): 35–54. http://dx.doi.org/10.14214/sf.30.
Hämet-Ahti L. (1963). Zonation of the mountain birch forest in northernmost Fennoscandia. Annales Botanici Fennici 34(4). 127 p.
Hänninen H. (1991). Does climatic warming increase the risk of frost damage in northern trees? Plant, Cell & Environment 5: 449–454.
Heikkinen O., Tuovinen M., Autio J. (2002). What determines the timberline? Fennia 180: 67–74.
Heinonen J. (1994). Koealojen puu- ja puustotunnusten laskentaohjelma KPL. Käyttöohje. Metsäntutkimuslaitoksen tiedonantoja - The Finnish Forest Research Institute, Research Papers 504. 80 p. ISBN 951-40-1369-7.
Hilli A., Hokkanen T., Hyvönen J., Sutinen M.-L. (2008). Long-term variation in Scots pine seed crop size and quality in northern Finland. Scandinavian Journal of Forest Research 23(5): 395–403. http://dx.doi.org/ 10.1080/02827580802334217.
Høgda K.A., Tømmervik H., Karlsen S.R. (2013). Trends in the start of the growing season in Fennoscandia 1982–2011. Remote Sensing 5: 4304–4318. http://dx.doi.org/10.3390/rs5094304.
Holtmeier F.-K. (2005). Change in the timberline ecotone in northern Finnish Lapland during the last thirty years. Reports from the Kevo Subarctic Research Station 23: 97–113.
Holtmeier F.-K., Broll G. (2011). Response of Scots Pine (Pinus sylvestris) to warming climate at its altitudinal limit in Northernmost Subarctic Finland. Arctic 64(3): 269–280. http://dx.doi.org/10.14430/arctic4118.
Hustich I. (1948). The Scots pine in northernmost Finland and its dependence on the climate in the last decades. Acta Botanica Fennica 42: 1–75.
Hyppönen M., Varmola M., Juntunen V., Lohl T., Mikkola K., Mäkitalo K., Timonen M. (2003). Metsien uudistuminen suojametsäalueella. Metsätieteen aikakauskirja 2003(1): 33–47.
Hyppönen M., Varmola M., Mikkola K., Jalkanen R. (2013). Metsien uudistaminen Lapin suojametsäalueella ja Pohjois-Suomen korkeilla mailla. In: Hyppönen M., Salminen S. (eds.). Metsänuudistaminen pohjoisen erityisolosuhteissa. Metlan työraportteja / Working Papers of the Finnish Forest Research Institute 255: 59–65. ISBN 978-951-40-2409-2.
Jalkanen R. (2003). Havupuutaimikoiden tuhojen esiintyminen ja merkittävyys Suomessa. Metsätieteen aikakauskirja 1/2003: 59–68.
Jalkanen R., Konôpka B. (1998). Snow-packing as a potential harmful factor on Picea abies, Pinus sylvestris and Betula pubescens at high altitude in northern Finland. European Journal of Forest Pathology 28: 373–382.
Jalkanen R., Aalto T., Pensa M., Salminen H. (2007). Development of Scots pine in the changing environment of the northern boreal zone in Finland. In: Taulavuori E., Taulavuori K. (eds.). Physiology of northern plants under changing environment. Research Signpost. p. 271–289.
Juntunen V., Neuvonen S. (2006). Natural Regeneration of Scots Pine and Norway Spruce close to the timberline in northern Finland. Silva Fennica 40(3): 443–458. http://dx.doi.org/10.14214/sf.329.
Juntunen V., Neuvonen S., Norokorpi Y., Tasanen T. (2002). Potential for timberline advance in northern Finland, as revealed by monitoring during 1983–99. Arctic 55(4): 348–361. http://dx.doi.org/10.14430/arctic719.
Kallio P., Hurme H., Eurola S., Norokorpi Y., Sepponen P. (1986). Research activities on the forest line in northern Finland. Arctic 39(1): 52–58.
Karlsen S.R., Elvebakk A., Johansen B. (2005). A vegetation-based method to map climatic variation in the arctic-boreal transition area of Finnmark, north-easternmost Norway. Journal of Biogeography 2005: 1161–1186. http://dx.doi.org/ 10.1111/j.1365-2699.2004.01199.x.
Kauppi P.E., Posch M., Pirinen P. (2014). Large impacts of climatic warming on growth of boreal forests since 1960. PLoS ONE 9(11): 1–6. http://dx.doi.org/ 10.1371/journal.pone.0111340.
Klein Tank A.M.G., Wijngaard J.B., Können G.P., Böhm R., Demarée G., Gocheva A., Mileta M., Pashiardis S., Hejkrlik L., Kern-Hansen C., Heino R., Bessemoulin P., Müller-Westermeier G., Tzanakou M., Szalai S., Pálsdóttir T., Fitzgerald D., Rubin S., Capaldo M., Maugeri M., Leitass A., Bukantis A., Aberfeld R., Van Engelen A.F.V., Forland E., Mietus M., Coelho F., Mares C., Razuvaev V., Nieplova E., Cegnar T., López J.A., Dahlström B., Moberg A., Kirchhofer W., Ceylan A., Pachaliuk O., Alexander L.V., Petrovic P. (2002). Daily dataset of 20th-century surface air temperature and precipitation series for the European Climate Assessment. International Journal of Climatology 22: 1441–1453. http://www.ecad.eu. [Cited 09 June 2015].
Kultti S., Mikkola K., Virtanen T., Timonen M., Eronen M. (2006). Past changes in the Scots pine forest line and climate in Finnish Lapland: a study based on mega fossils, lake sediments, and GIS-based vegetation and climate data. Holocene 16(3): 381–391. http://dx.doi.org/ 10.1191/0959683606hl934rp.
Lehtonen I., Hoppula P., Pirinen P., Gregow H. (2014). Modelling crown snow loads in Finland: a comparison of two methods. Silva Fennica 48(3): 1–30. http://dx.doi.org/10.14214/sf.1120.
Leinonen I. (1995). A simulation model for the annual frost hardiness and freeze damage of Scots pine. Annals of Botany 78: 687–693.
Lenth R.V. (2014). lsmeans: Least-Squares Means. R package version 2.05. http://CRAN.R-project.org/package=lsmeans. [Cited 18 Oct 2014].
Linkosalo T., Häkkinen R., Terhivuo J., Tuomenvirta H., Hari P. (2009). The time series of flowering and leaf bud burst of boreal trees (1846-2005) support the direct temperature observations of climatic warming. Agricultural and forest meteorology 149: 453–461. http://dx.doi.org/10.1016/j.agrformet.2008.09.006.
Marchand P. (1987). Life in cold: an introduction to winter ecology. University Press of New England. 320 p.
Mattila E. (2014a). Ylä-Lapin talvilaidunarvioinnin tuloksia – Uusimmat arviot vuodelta 2012 ja vastaavia tuloksia vuodelta 2004. Metlan työraportteja / Working Papers of the Finnish Forest Research Institute 282. 55 p.
Mattila E. (2014b). Porojen talvilaitumien määrä ja laatu poronhoitoalueen etelä- ja keskiosissa – Uudet arviot vuosilta 2009–2013 ja keskeisten laiduntunnusten kehitys osin 1970-luvulta lähtien. Metlan työraportteja / Working Papers of the Finnish Forest Research Institute 304.
Mikkonen S., Laine M., Mäkelä H.M., Gregow H., Tuomenvirta H., Lahtinen M., Laaksonen A. (2014). Trends in the average temperature in Finland, 1847-2013. In: Christakos G. (ed.). Stochastic environmental research and risk assessment. Springer-Verlag, Berlin-Heidelberg. p. 1–9. http://dx.doi.org/ 10.1007/s00477-014-0992-2.
Mustajärvi K., Merilä P., Derome J., Lindroos A.-J., Helmisaari H.-S., Nöjd P., Ukonmaanaho L. (2008). Fluxes of dissolved organic and inorganic nitrogen in relation to stand characteristics and latitude in Scots pine and Norway spruce stands in Finland. Boreal Environment Research 13(suppl. B): 3–21.
Neuvonen S., Niemelä P., Virtanen T. (1999). Climatic change and insect outbreaks in boreal forests: the role of winter temperatures. Ecological Bulletins 47: 63–67.
Niemelä P., Rousi M., Saarenmaa H. (1987). Topographical delimitation of Neodiprion sertifer (Hym., Diprionidae) outbreaks on Scots pine in relation to needle quality. Journal of Applied Entomology 103: 84–91.
Ojansuu R., Henttonen H. (1983). Estimation of local values of monthly mean temperature, effective temperature sum and precipitation sum from the measurements made by the Finnish Meteorological Office. Silva Fennica 17: 143–160.
Pinheiro J., Bates D., DebRoy S., Sarkar D., the R Core team (2013). nlme: Linear and nonlinear mixed effects models. R package version 3: 1–113.
R Core Team (2013). R: a language and environment for statistical computing. R. Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org/. [Cited 18 Oct 2014].
Repo T., Hänninen H., Kellomäki S. (1996). The effects of long-term elevation of air temperature and CO2 on the frost hardiness of Scots pine. Plant, Cell & Environment 19: 209–216.
Salminen H., Jalkanen R. (2015). Modelling of bud break of Scots pine in northern Finland in 1908–2014. Frontiers in Plant Science 6: 104. http://dx.doi.org/10.3389%2Ffpls.2015.00104.
Seo J.-W., Salminen H., Jalkanen R., Eckstein D. (2010). Chronological coherence between intra-annual height and radial growth of Scots Pine (Pinus sylvestris L.) in the northern boreal zone of Finland. Baltic Forestry 16(1): 57–65.
Seppä H., Nyman M., Korhola A., Weckström J. (2002). Changes of treelines and alpine vegetation in relation to post-glacial climate dynamics in northern Fennoscandia based on pollen and chironomid records. Journal of Quaternary Science 17(4): 287–301. http://dx.doi.org/ 10.1002/jqs.678.
Snowdon P. (1991). A ratio estimator for bias correction in logarithmic regression. Canadian Journal of Forest Research 21: 720–724.
Sutinen R., Närhi P., Middleton M., Hänninen P., Timonen M., Sutinen M.-L. (2012). Advance of Norway spruce (Picea abies) onto mafic Lommoltunturi fell in Finnish Lapland during the last 200 years. Boreas 41: 367–378. http://dx.doi.org/ 10.1111/j.1502-3885.2011.00238.x.
Tietäväinen H., Tuomenvirta H., Venäläinen A. (2010). Annual and seasonal mean temperatures in Finland during the last 160 years based on gridded temperature data. International Journal of Climatology 30: 2247–2256. http://dx.doi.org/ 10.1002/joc.2046.
Tuhkanen S. (1980). Climatic parameters and indices in plant geography. Acta Phytogeographica Suecica 67: 1–110.
Vajda A., Venäläinen A. (2005). Feedback processes between climate, surface and vegetation at the northern climatological tree line (Finnish Lapland). Boreal Environment Research 10: 299–314.
Väre H., Ohtonen R., Mikkola K. (1996). The effect and extent of heavy grazing by reindeer in oligotrophic pine heaths in northeastern Fennoscandia. Ecography 19: 245–253.
Veteli T.O., Lahtinen A., Repo T., Niemelä P., Varama M. (2005). Geographic variation in winter freezing susceptibility in the eggs of the European pine sawfly (Neodiprion sertifer). Agricultural and Forest Entomology 7: 115–120. http://dx.doi.org/ 10.1111/j.1461-9555.2005.00259.x.
Virtanen T., Neuvonen S., Nikula A., Varama M., Niemelä P. (1996). Climate change and the risks of Neodiprion sertifer outbreaks on Scots pine. Silva Fennica 30(2–3): 169–177.
Total of 55 references
