Markus Melin  ,
Tiina Ylioja,
Leena Aarnio,
Katri Hamunen,
Seppo Nevalainen,
Antti Pouttu,
Heli Viiri
,
Tiina Ylioja,
Leena Aarnio,
Katri Hamunen,
Seppo Nevalainen,
Antti Pouttu,
Heli Viiri
Emergence levels of pine shoot beetles from roundwood piles of Scots pine and the cascading damage in the surrounding forests
Melin M., Ylioja T., Aarnio L., Hamunen K., Nevalainen S., Pouttu A., Viiri H. (2021). Emergence levels of pine shoot beetles from roundwood piles of Scots pine and the cascading damage in the surrounding forests. Silva Fennica vol. 55 no. 5 article id 10525. https://doi.org/10.14214/sf.10525
Highlights
- Emerged pine shoot beetles were counted from piles of harvested Scots pine, and the shoot damage in the surrounding forests was measured
- Damage was noticeable up to a distance of 40–60 m, being more severe near large piles
- For piles smaller than 50 m3 the level of damage (fallen shoots) was mainly below known thresholds for growth losses
- Logs with harvester-damaged bark were significantly less colonized by the beetles.
Abstract
Bark beetles are amongst the most aggressive pest agents of coniferous forests. Due to this, many boreal countries have designated laws aiming to lower the risk of bark beetle epidemics. Finland’s forest legislation has pre-emptive measures targeted against bark beetles, and for Scots pine (Pinus sylvestris L.), the law concerns pine shoot beetles (Tomicus spp.). This study used data collected around 25 piles of Scots pine roundwood that were harvested in the winter but left in the forest until the following November. Thus, the pine shoot beetles were able to use the piles for breeding. We assessed the number of emerged insects from the piles and the cascading damage they caused in the surrounding forests. All roundwood piles, regardless of their volume, were used by the beetles for breeding. Highest densities of beetle exit holes were found from the parts of the log with thick and intact bark. If the bark of the log was damaged by the harvester head, the number of beetles decreased significantly. Depending on the volume of the roundwood pile, the cascading damage (fallen shoots) was noticeable up to ca. 40–60 m from the roundwood pile. Storing of piles smaller than 50 m3 did not cause excess damage. The number of fallen shoots per tree was generally below the known thresholds for when growth losses can occur. However, the study was conducted in mature forests, and it can be assumed that the recorded damage levels would severely affect the growth of young pines, raising the question of where to store the roundwood. As with other bark beetles, the role of Tomicus beetles as damage agents may change in the future, but based on this as well as past studies, the species can be viewed as a notable damage agents only around long-term wood storage sites in the current northern conditions.
Keywords
forest management;
forestry;
Tomicus piniperda;
Tomicus minor;
roundwood;
legislation;
timber storage
-
Melin,
Natural Resources Institute Finland (Luke), Bioeconomy and environment, Yliopistokatu 6b, FI-80100 Joensuu, Finland
E-mail
markus.melin@luke.fi
- Ylioja, Natural Resources Institute Finland (Luke), Natural resources, Latokartanonkaari 9, FI-00790 Helsinki, Finland E-mail tiina.ylioja@luke.fi
- Aarnio, Natural Resources Institute Finland (Luke), Natural resources, Latokartanonkaari 9, FI-00790 Helsinki, Finland E-mail leena.aarnio@luke.fi
- Hamunen, Natural Resources Institute Finland (Luke), Bioeconomy and environment, Yliopistokatu 6b, FI-80100 Joensuu, Finland E-mail katri.hamunen@luke.fi
- Nevalainen, Natural Resources Institute Finland (Luke), Bioeconomy and environment, Yliopistokatu 6b, FI-80100 Joensuu, Finland E-mail seppo.nevalainen@gmail.com
- Pouttu, Natural Resources Institute Finland (Luke), Natural resources, Latokartanonkaari 9, FI-00790 Helsinki, Finland E-mail antti.pouttu@kolumbus.fi
- Viiri, Natural Resources Institute Finland (Luke), Bioeconomy and environment, Yliopistokatu 6b, FI-80100 Joensuu, Finland; UPM-Kymmene Oyj, UPM Forest, Peltokatu 26 C, PL 85, FI-33100, Tampere, Finland E-mail heli.viiri@upm.com
Received 15 February 2021 Accepted 28 October 2021 Published 2 November 2021
Views 47192
Available at https://doi.org/10.14214/sf.10525 | Download PDF

1 Introduction
Countries where forestry is an important industry typically have designated laws aiming to safeguard its sustainability. These laws can target forest operations, ownership and management as well as how these relate to forest health. The latter, especially in the boreal zone, is becoming increasingly important as the most severe damage agents are bark beetles (Coleoptera: Curculionidae, Scolytinae) – a group of insects known for their capability to cause large-scale tree mortality and benefiting from the predicted climatic changes (Seidl and Rammer 2017). Therefore, it is no surprise that the functioning of the current laws is re-evaluated on a regular basis.
In Finland, the Forest Damages Prevention Act (Finlex 2013) emphasises precautionary measures to maintain bark beetle populations at endemic levels, and the functioning of this law has recently been evaluated (Kniivilä et al. 2020), which also involved assessing the regulations targeted against bark beetles. So far, Finland’s forests have not experienced severe bark beetle epidemics. The species that the legislation focuses on are the spruce bark beetle (Ips typographus L.) and the common- and lesser pine shoot beetle (Tomicus piniperda L., Tomicus minor L.). For the pine shoot beetles, the legislation mandates that all roundwood harvested or fallen (e.g., due to wind) between September 1 and May 31 must be transported from the forest before the assumed emergence of the beetle’s first generation, which happens between June and July, depending on the area. This regulation concerns freshly cut or fallen Scots pine (Pinus sylvestris L.) roundwood when the volume exceeds 20 m3 ha–1. The justification is that by removing the wood early enough, the beetle larvae that are growing under the bark cannot develop into adult beetles and emerge into the forests, inhibiting their potential damage to the surrounding forest. However, these regulations have not been tested in the field during the machine-based forest harvesting age – a point also raised in the previous evaluation of the current forest damage legislation (Kniivilä et al. 2020).
The common pine shoot beetle (hereafter referred to as “pine shoot beetle”) and the lesser pine shoot beetle are the bark beetles responsible for most bark beetle damage on Finnish pines. The beetle species are among the first bark beetles to start breeding after the winter when the temperatures reach 10–12 °C, which, in Southern and Central Finland, can be anywhere between late-March and late-April, depending on the year. Breeding takes place under the tree bark, in the phloem, and after successful breeding, the female creates a gallery under the bark along which it lays the eggs. Tomicus piniperda typically uses the phloem under thick bark and T. minor that under thinner bark (Långström 1984). While T. piniperda colonises logs as breeding material, T. minor seldom does so (Juutinen 1978). Juvenile beetles typically emerge during June–July. Both the adults and the emerged juvenile beetles attack the surrounding forests, where they feed in the shoots of the trees, typically in the tallest canopies. As with other bark beetles, both pine shoot beetles also transport blue-stain fungi with them as they enter the tree phloem (Solheim and Långström 1991; Silva 2011). The damage they cause is thus affecting the tree in more than one way: galleries in the phloem, blue-stain fungi transported to the phloem and shoot damage in the canopy. When the populations are abundant, the damage is distinguishable: treetops deprived of shoots and high amounts of shoots fallen to the ground (Fig. 1).

Fig. 1. Top-left and centre: damage by pine shoot beetle results in pine trees deprived of their shoots. Bottom-left: shoots fallen to the ground. Right: two vertical monogamous galleries created by pine shoot beetle females, with larval galleries breaking into adjacent directions. Images by Antti Pouttu and Markus Melin, Natural Resources Institute Finland (CC-BY SA 4.0).
Shoot losses, although severe, do not commonly result in the death of the tree. Nilsson (1974) estimated that significant growth losses in mature pines would occur after a tree has lost 100–150 shoots, but in the case of young pines, notable growth losses can occur after the tree has lost 15–20 shoots. Andersson (1973) recorded growth losses of 19% already at the level of 30 fallen shoots. Ericsson et al. (1985) removed 200 shoots from mature pines but recorded only minor growth losses: on average, the growth losses regarding tree volume were 16–20% and lasted for 2 years. In more recent papers, the level of pine shoot beetle damage has been estimated following storms after which high amounts of fallen pines have been available as breeding material for the beetles. In Finland, Komonen et al. (2009) quantified damage at the level of 54–165 fallen shoots per tree after a storm felled large amounts of mature pines in an island. Annila et al. (1999) studied Tomicus attacks on Scots pines that had suffered defoliation by the common pine sawfly (Diprion pini L.) and concluded that even after heavy defoliation by D. pini, the pines were still able to defend themselves against Tomicus attacks to the point that no mortality was recorded. In Sweden, Öhrn et al. (2018) documented Tomicus-induced tree-wise shoot losses at a level of ca. 30–100 shoots per tree following a storm. Despite many papers documenting growth losses, none of the Fennoscandian researchers have documented tree deaths where Tomicus beetles would have been the primary cause.
A question that, to a large extent, is also unanswered is the effect of forestry operations, especially that of stacks of roundwood that are temporarily stored in the forests (Fig. 2). These kinds of piles are formed on a regular basis throughout the country and their removal is what the Finnish Forest Damages Prevention Act directly requires.

Fig. 2. A common sight in Finland: pile of roundwood (here, Scots pine) harvested during the previous winter is waiting to be transported to the factory. Image by Markus Melin, Natural Resources Institute Finland (CC-BY SA 4.0).
How the kinds of piles illustrated in Fig. 2 affect the abundance of the pine shoot beetle in the surrounding forests is a critical question since harvesting is done on a continuous basis throughout the country and throughout the year. After all, in the mentioned re-evaluation of the Forest Damages Prevention Act (Kniivilä et al. 2020), researchers concluded that the current dates by which the timber must be transported from the forest are, to a large extent, invalid in the current climate: the pine shoot beetles can emerge before the set deadlines. In addition, it is known that freshly cut roundwood (as the one illustrated in Fig. 2) is commonly used by the pine-shoot beetles for breeding (Borkowski and Skrzecz 2016). Also, in re-capture studies, the majority of emerged pine shoot beetles attacked a tree within 400 meters from where they had emerged (Barak et al. 2000). Together, these results suggest that a stored roundwood pile such as the one depicted in Fig. 2 is likely to increase the risk of pine shoot beetle damage in the surrounding forests.
In this context, our study assessed 1) the level of shoot damage (and potential growth losses) to the surrounding forest caused by roundwood piles that are not removed by the legislation date; 2) how the risk relates to the volume of the roundwood pile; and 3) how far away from the pile the damage can spread. We also estimated the number of beetles emerging from the piles and how this relates to the diameter of the log and the properties of its bark.
2 Materials and methods
2.1 Study area and field measurements
The study was conducted across Central- and Eastern Finland in mature, pine-dominated forests surrounding 25 roundwood piles with varying volumes (Fig. 3). The piles were created with standard forestry machines and operated with no special treatment. The trees were cut in the winter (February–March) preceding the field measurements, meaning that they were in place and usable for the pine shoot beetles when they begin to breed in April–May. The piles were purposely stored in the field beyond the dates of wood removal set in the Forest Damages Prevention Act (Finlex 2013); this breach of forest legislation was done with permission from the Finnish Forest Centre. Six of the piles were measured in 2017 (between 27 Oct. 19 Nov.) and 19 in 2020 (between 29 Aug. and 30 Oct.). Unfortunately, we were not able to affect the spatial coverage of the data: it was entirely up to the companies to decide which piles they were able to spare for our research purposes and from which parts of the country. We accepted this and conducted the field work around all the piles that were offered to us.
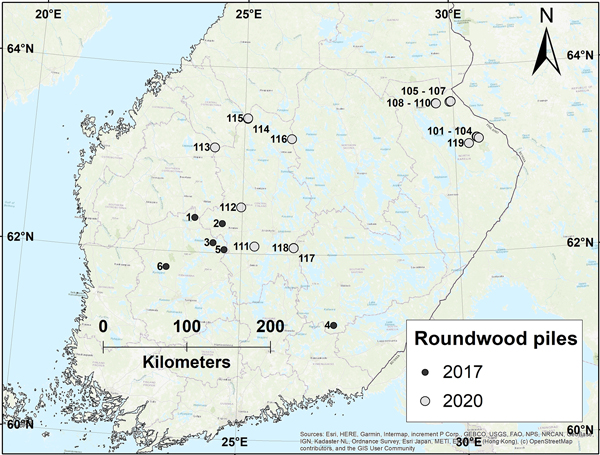
Fig. 3. Location of the roundwood piles used in the study on pine shoot beetle damage in the surrounding forests, separated by the year of field measurements.
The field work consisted of measuring 1) the volume of the roundwood pile; 2) the number of beetles that emerged from them; and 3) the shoot damage they caused in the surrounding forests. Volumes of the roundwood piles were measured with common methods certified by the Ministry of Agriculture and Forestry (Metsäteho 2003). Harvester-measured volume was attainable for one pile only (Pile 119). The manual methods used for the other piles involved the following steps: the piles were split into two-meter wide sections, and the dimensions (height, width, length) of each section was measured individually. After this, the section-wise dimensions were used to calculate the section-wise gross volumes, and these were then combined to attain the volume of the entire pile. The gross volume values were then adjusted with fixed conversion factors (Metsäteho 2003) that were based on 1) how many branches the logs of the pile contained, 2) how bent/crooked the logs of the pile were, 3) how densely the logs were stacked in the pile, and 4) the average diameter of the logs in the pile. Variables 1–3 were assessed visually as averages across the entire pile, on a scale from 1 to 5. The average diameter was estimated by measuring the diameters of log heads from two samples placed at 25th and 75th quantiles in relation to the width of the pile. For example, for a 20-meter wide pile, these samples were taken 5 m away from both ends of the pile. At these sites, the log head diameters were measured with callipers from a circular area with a 60 cm diameter. These diameter measurements were then averaged to attain the pile-wise average diameter.
The number of emerged beetles was estimated by counting their exit holes from individual logs from the topmost layer of the piles. The sampled logs were selected at 2-meter intervals so that the first log was selected 2 m from the end of the pile, the next one 2 m from the previous one etc. Emphasis was placed on assuring that logs of different diameters were present in the data: if, for example, two or three logs intersected at the 2-m mark, the log having a diameter not present in the data already was selected. The pine shoot beetle emergence holes were counted from 10 × 30-cm rectangular sampling areas. From each selected log, sampling on an area of 10 × 30 cm was done at 1-meter intervals: the first sample was taken 50 cm away from the base of the log, the next one 100 cm from the previous one, etc., pending that the last sample was at least 50 cm away from the tip of the log. Holes made by other beetles, such as Pityogenes spp. and Trypodendron lineatum, were ignored. For accuracy, the samples from where the exit holes were calculated were taken with a 10 × 30-cm frame, which was placed on the top of the log, at the desired spot. The diameter (in cm) of the log at the centre of each 10 × 30-cm sampling site was also measured, as was the proportion of non-intact bark (damaged by the harvester head) that would be unusable for the pine shoot beetle. Log diameters were measured with callipers, and the proportion of harvester-damaged bark was estimated visually from the sample area as a percentage of damaged area from the total sample area (in 10% classes). In the 2020 field measurements, the bark at each sample site was also categorised in three categories based on its thickness: thin, medium, thick. This estimation was based on visual properties of the bark (Fig. 4).

Fig. 4. The three categories of bark thickness used in the 2020 field survey when counting the number of beetle exit holes from the logs; the bark of Scots pine is changing from thick to thin when moving from the base of a log (left) into its top (right).
Altogether, the data for assessing the emergence levels of the beetles consisted of 544 exit hole samples taken from 155 different logs. Due to weather conditions (heavy rain, wind and limited visibility), the sampling of exit holes was not conducted on four of the piles in the 2020 survey as conducting of the full measurements on top of the wet and slippery piles was deemed too hazardous. Yet, the presence of Tomicus exit holes was still visually verified also for these piles.
The damage that the emerged pine shoot beetles caused in the surrounding forests was estimated by counting the fallen shoots (Fig. 1, bottom-left corner) from circular sample plots with a 1.78-m radius (ca. 10 m2). This method has been proven to be more error-free than other methods based on e.g. crown properties (Borkowski 2007), and it was also considerably faster than any crown-based measurements would have been. Shoots fallen during earlier years were omitted: they were detectable based on colour, decay and, in some cases, on the presence of needle blight. The counting of fallen shoots was started so that from each roundwood pile, at least two line transects were drawn, in two perpendicular directions. The circular plots where the shoots were counted were then placed along these transects at 10-meter intervals so that the first plot was placed as near as possible to the roundwood pile, but still within a forest and not on a forest road, for instance. New plots were placed along the transect lines until the level of fallen shoots had dropped into a “baseline” level of pine shoot beetle damage: one fallen shoot per m2 (Komonen et al. 2009). Transect lengths varied between 44 and 100 meters, with an average of 61 meters, measured from the centre of the pile. By default, two transects were drawn into two perpendicular directions from the pile, but if the pile was surrounded by distinctly different forest stands (e.g., a mature pine stand and a young pine seedling stand), the transects were drawn into the different stands regardless of their bearing, to account for this variation. Every plot along every transect was marked in the field with a small (1 × 50 cm), blue-painted wooden pole, which was placed at the centre of the plot. The poles were left in the field for potential future measurements.
In the 2020 field measurements, the structure of the surrounding forests was also estimated by measuring average height, diameter and basal area from one of the field plots in each of the transects. The plot where these measurements were taken was selected based on how well it represented the overall forest structure at the site; most often, this was the middle plot of one of the transects. The purpose of this measurement was to obtain an estimate about the density of the forest, which was later used in estimating the average tree-wise shoot losses. In addition, the presence of alternative factors that may have affected the abundance of pine shoot beetles (presence of snow- and wind damaged trees, logging residues) was also recorded. Altogether, 222 field plots were measured for estimating the shoot damage caused by the beetles (Table 1).
| Table 1. Auxiliary information on the roundwood piles used in the study on pine shoot beetle damage in forests surrounding the piles. The coordinates are in the WGS84 system (EPSG: 4326). | ||||||
| Pile | Latitude | Longitude | Year | Volume (m3) | No. of measured field plots (shoot damage) | No. of measured samples (emerged insects) |
| 1 | 23.9025844 | 62.3706153 | 2017 | 356 | 9 | 20 |
| 2 | 24.5415178 | 62.3168368 | 2017 | 331 | 7 | 30 |
| 3 | 24.3453392 | 62.108855 | 2017 | 34 | 7 | 20 |
| 4 | 27.1107396 | 61.2459632 | 2017 | 42 | 5 | 16 |
| 5 | 24.6045835 | 62.0375501 | 2017 | 42 | 8 | 16 |
| 6 | 23.3038068 | 61.8281155 | 2017 | 130 | 6 | 31 |
| 101 | 30.5765414 | 63.2180891 | 2020 | 145 | 13 | 29 |
| 102 | 30.5371486 | 63.2287643 | 2020 | 127 | 8 | 29 |
| 103 | 30.5314965 | 63.22911 | 2020 | 59 | 10 | 17 |
| 104 | 30.5243809 | 63.2310969 | 2020 | 149 | 9 | 32 |
| 105 | 29.9444647 | 63.6232792 | 2020 | 40 | 9 | 13 |
| 106 | 29.9426211 | 63.6244717 | 2020 | 39 | 8 | 10 |
| 107 | 29.9159022 | 63.6183939 | 2020 | 144 | 8 | 33 |
| 108 | 29.5883251 | 63.60701 | 2020 | 125 | 8 | 0 |
| 109 | 29.5854189 | 63.6088174 | 2020 | 64 | 9 | 0 |
| 110 | 29.5818343 | 63.6096894 | 2020 | 124 | 8 | 34 |
| 111 | 25.2972062 | 62.0822797 | 2020 | 173 | 12 | 36 |
| 112 | 24.9662172 | 62.4973184 | 2020 | 110 | 13 | 28 |
| 113 | 24.3008883 | 63.1313398 | 2020 | 131 | 9 | 34 |
| 114 | 25.0644776 | 63.4503408 | 2020 | 297 | 8 | 0 |
| 115 | 25.0680649 | 63.4565652 | 2020 | 138 | 8 | 27 |
| 116 | 26.1272094 | 63.2416311 | 2020 | 242 | 9 | 0 |
| 117 | 26.1974721 | 62.0754757 | 2020 | 55 | 9 | 21 |
| 118 | 26.1973003 | 62.0728962 | 2020 | 175 | 9 | 42 |
| 119 | 30.3350948 | 63.1643917 | 2020 | 151 | 13 | 34 |
2.2 Statistical analysis
The statistical analysis focused on testing and quantifying the extent to which 1) the roundwood piles were used for breeding by the pine shoot beetles, and 2) the storage of the roundwood piles led to cascading damage in the surrounding forests in the form of fallen shoots. For these, two linear mixed effects models were built. Linear Mixed Effects models (LME) constitute of a fixed part (the predictors) and a random part that accounts for potential grouping of the data; here, the data were clustered according to the individual roundwood piles that were measured over two years. This nested structure was accounted for with the use of random effects for ‘Year’ and for ‘Pile’, with the latter being nested within the former.
The first LME model focused on the emerged pine shoot beetles that were counted from the 10 × 30-cm samples. In this model, the number of exit holes per m2 was the response variable and the fixed predictors were log diameter, bark thickness, the proportion of damaged bark, the basal area of pine in the surrounding forest, site type and the presence of damaged pines or logging residues. The latter three were included as they may affect the general level of pine shoot beetles in the area, regardless of the roundwood pile itself. The second model focused on the number of fallen shoots, namely the pine shoot beetle damage. Here, the number of fallen shoots per plot was the response variable, whereas the fixed predictors were the distance between the plot and the roundwood pile, the volume of the roundwood pile, the basal area of pine in the surrounding forest, the site type and the presence of damaged pines or logging residues. Both models followed the simplest form of an LME model, which can be expressed as follows:
![]()
where X is the design matrix for fixed effects and β is the vector for fixed effects, Z is the design matrix for random effects and b is the vector for random effects, whereas ε is the vector for observation errors (Pinheiro and Bates 2004). As stated, both models included two random effect terms for ‘Year’ and for ‘Pile’, meaning that the variation caused by a different survey year or differences between the piles were accounted for. Variable selection in both models was done by backwards selection, where the least significant variable was dropped, and the model was re-built. The process was then iterated until only significant (p < 0.05) variables remained in the model. Table 2 lists the candidate predictor variables as well as the final ones for both models. In addition to modelling, the numbers of the emerged insects and dropped shoots were also analysed and visualised with basic descriptive- and summary statistics. All data analyses, statistical analyses and visualisations of the results were conducted in R (R Core Team 2021), using the packages ‘ggplot2’ (Wickham 2016) for graphics and ‘nlme’ (Pinheiro et al. 2020) for the mixed-effects modelling.
| Table 2. Candidate variables in the LME models on pine shoot beetle damage in forests close to roundwood piles and on the number of emerged insects from the roundwood piles. * indicates a variable that was retained in the final model. | ||
| Model | Variable | Description |
| Model 1: Emerged insects | Emerged insects (the response) | Number of pine shoot beetle emergence holes in the sampling area, holes per m2. |
| Log diameter* | Diameter of the log (in centimetres), measured from the middle of the sampling window. | |
| Bark thickness* | Thickness of the bark, three categories: thin, medium, thick. | |
| Bark damage* | Proportion of harvester-damaged bark, estimated from the sampling areas at 10% intervals on a 0–100% scale. In the modelling, the variables were tested also as a factorial variable with five levels (0–20% bark damage, 20–40% bark damage etc.) | |
| Basal area, pine | Basal area of the pine in the surrounding forests, measured from the field plots, m2 ha–1. | |
| Site type | Site type of the surrounding forest according to Cajander (1949), estimated from the field plots. | |
| Damaged pine | Occurrence of pine shoot beetle breeding material in the surroundings (binary): snow or wind damage, logging residues, etc. | |
| Model 2: Fallen shoots | Shoot damage (the response) | Number of fallen shoots in a field plot, shoots per m2. |
| Distance* | Distance (in meters) between the plot to the roundwood pile. | |
| Volume* | Volume of the neighbouring roundwood pile. | |
| Basal area, pine | As described above. | |
| Site type | As described above. | |
| Damaged pine | As described above. | |
3 Results
3.1 Emergence of insects from the roundwood piles
All studied piles were used for reproduction by the pine shoot beetles, which was evident based on the number of emerged beetles. Nearly all exit holes were made by T. piniperda, and exit holes made by T. minor were noticed that rarely that they were not even quantified. The main factors behind the number of emerged beetles were the diameter of the log, the thickness of the bark and the proportion of bark damaged by the harvester. The proportion of bark damaged by the harvester had a more significant effect on the exit holes when modelled as a factorial predictor in 20% classes (Table 3).
| Table 3. The LME model on the number of emerged beetles from the forest-stored roundwood piles. | |||
| FIXED EFFECTS | |||
| Parameter | Value | Std. error | p-value |
| Intercept | –103.94 | 73.48 | 0.158 |
| Bark, medium | –1.80 | 28.15 | 0.949 |
| Bark, thick | –117.83 | 32.77 | <0.001 |
| Bark damage, 20–40% | –64.74 | 37.50 | 0.0851 |
| Bark damage, 40–60% | –98.41 | 36.66 | 0.0076 |
| Bark damage, 60–80% | –159.06 | 58.26 | 0.0066 |
| Bark damage, 80–100% | –205.41 | 69.17 | 0.0032 |
| Log diameter | 23.92 | 3.41 | <0.001 |
| RANDOM EFFECTS | |||
| Std. dev. | Residual | ||
| Year | 29.4 | ||
| Roundwood pile | 80.79 | 223.25 | |
The larger the diameter of the log, the more beetles emerged from it. In a similar but opposite manner, the more the harvester head had damaged the bark, the less beetles emerged from it. This became especially clear when the proportion of harvested-damaged bark increased to over 40% (Table 3). The relationship between the percentage of bark damaged by the harvester and the number of beetle exit holes is separately illustrated in Fig. 5.
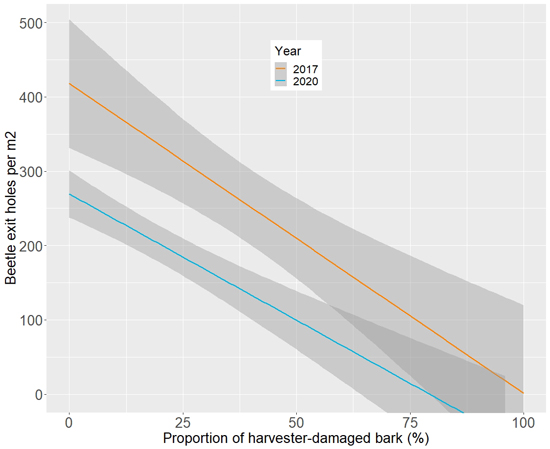
Fig. 5. Visualisation of the relationship between the variable BarkDamage (percentage of bark damaged by the harvester) and the number of beetle exit holes counted from the logs of the forest-stored roundwood piles. The shading illustrates the 95% confidence interval and the different colours indicate different survey years.
Regardless of the diameter, thin-barked logs contained the least amount of beetle emergence holes. Average values for thin-barked logs were 69 holes per m2 (range from 0to 667); for medium-barked logs, the average was 220 (range from 0to 1233), and for the logs with thick bark, the average was 296 (range from 0to 1333) (Table 4).
| Table 4. Summary statistics on the number of Tomicus piniperda exit holes from each roundwood pile. | |||
| Pile | No. of emergence holes/m2 | ||
| Average | Maximum | Std. dev. | |
| 1 | 162 | 1000 | 274 |
| 2 | 238 | 1063 | 273 |
| 3 | 592 | 1300 | 412 |
| 4 | 90 | 367 | 100 |
| 5 | 392 | 900 | 233 |
| 6 | 121 | 740 | 216 |
| 101 | 152 | 633 | 181 |
| 102 | 80 | 533 | 145 |
| 103 | 137 | 700 | 222 |
| 104 | 178 | 933 | 254 |
| 105 | 326 | 900 | 316 |
| 106 | 337 | 1100 | 297 |
| 107 | 226 | 900 | 270 |
| 110 | 303 | 1267 | 371 |
| 111 | 230 | 800 | 221 |
| 112 | 104 | 1333 | 254 |
| 113 | 192 | 900 | 255 |
| 115 | 264 | 900 | 287 |
| 117 | 138 | 633 | 183 |
| 118 | 233 | 1233 | 323 |
| 119 | 189 | 733 | 217 |
| By bark thickness* | Average | Maximum | Std. dev. |
| Thin bark | 69 | 667 | 156 |
| Medium bark | 220 | 1233 | 287 |
| Thick bark | 296 | 1333 | 275 |
| * Only for piles 101–119, year 2020 measurements | |||
3.2 Shoot damage in the surrounding forests
The emergence of large numbers of beetles resulted in shoot damage in the surrounding forests around each roundwood pile. The scale of the damage was driven mostly by the volume of the pile. Logging residues and wind-felled trees, if present, increased the level of damage. While the overall level of the shoot damage was higher around piles with the highest volumes, the damage, regardless of the pile’s volume, was always more severe in plots closest to the roundwood piles (Table 5). The variation between the individual piles was higher than that between the survey years (Table 5, random effects).
| Table 5. The LME model on the number of fallen shoots per plot (the damage) in a study on pine shoot beetle damage in forests close to roundwood piles. | |||
| FIXED EFFECTS | |||
| Parameter | Value | Std. error | p-value |
| Intercept | 12.76 | 10.75 | 0.24 |
| Distance to pile | –0.69 | 0.11 | <0.001 |
| Volume of pile | 0.21 | 0.06 | 0.003 |
| Residues | 15.60 | 6.10 | 0.01 |
| RANDOM EFFECTS | |||
| Std. dev. | Residual | ||
| Year | 4.63 | ||
| Roundwood pile | 14.84 | 23.3 | |
On average (depending on the volume of the pile), shoot damage ceased at ca. 40–60 m away from the roundwood pile: at these distances, the number of fallen shoots was close to the baseline level that can be assumed to be normal in boreal forests (Fig. 6).
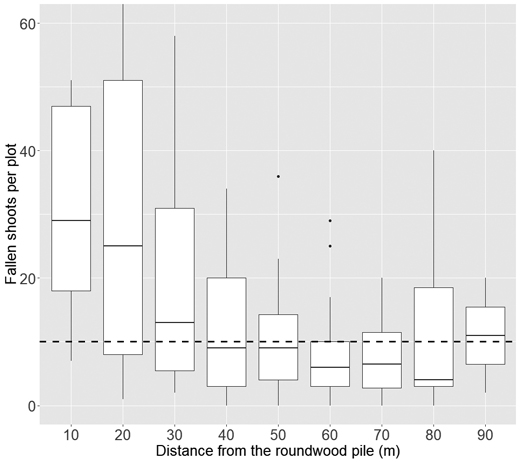
Fig. 6. Number of fallen shoots per plot at different distances from the roundwood piles. At each boxplot, the black horizontal line indicates the median and the white vertical bar the 25th and 75th quantiles. The dashed horizontal line represents the baseline level of fallen shoots that can be considered “normal” in boreal forests.
The effect of the pile volume on the number of fallen shoots in the surrounding forests is further illustrated in Fig. 7. For example, the damage around roundwood piles smaller than 50 m3 is close to the baseline level, whereas the damage around piles with larger volumes is clearly more severe in the plots closer to the pile.
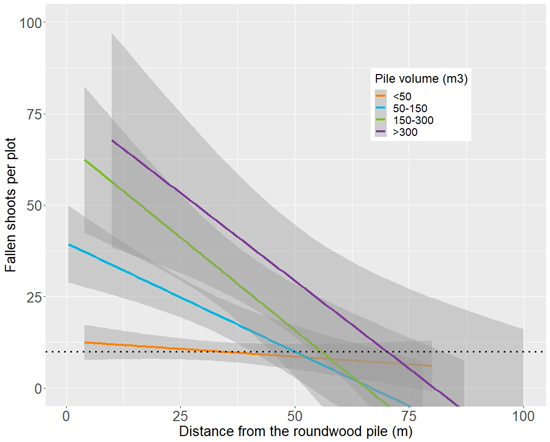
Fig. 7. Linear relationship between the distance from roundwood piles of varying volumes and the number of fallen shoots in a plot. For this illustration, the originally continuous variable Volume was classified in the depicted classes. The grey shading illustrates the 95% confidence intervals.
As the 2020 field measurements included the structure of forests surrounding each pile, the number of fallen shoots per plot was convertible into an approximation about the number of fallen shoots per tree, which varied between 0and 517 fallen shoots per tree. Maximum numbers (top three) were recorded in plots surrounding piles 110 (517 fallen shoots per tree), 114 (247 fallen shoots) and 109 (197 fallen shoots per tree). Altogether, tree-wise losses higher than 100 fallen shoots per tree were exceeded around piles 109, 110, 111, 112, 114 and 116. Of these, piles 109, 110, 112 and 116 had logging residues in the forests surrounding them, which had an increasing effect on the level of shoot damage (Table 6).
| Table 6. Tree-level damage (fallen shoots) caused by pine shoot beetles in relation to the distance from the pile. | ||||
| Distance from the pile (m) | Fallen shoots per tree | |||
| Min | Max | Mean | Std. dev. | |
| <10 | 9 | 197 | 47 | 50 |
| 10–20 | 1 | 517 | 63 | 100 |
| 20–30 | 2 | 247 | 40 | 45 |
| 30–40 | 0 | 171 | 23 | 25 |
| 40–50 | 0 | 92 | 23 | 25 |
| 50–60 | 0 | 74 | 19 | 22 |
| 60–70 | 0 | 46 | 18 | 20 |
| >70 | 0 | 100 | 41 | 50 |
4 Discussion
In this study, we assessed the extent to which temporarily stored roundwood piles of Scots pine result in pine shoot beetle damage in the surrounding forests – if not removed as the current Finnish Forest Damage Prevention Act requires. The piles were standard piles in the sense that such piles (as displayed in Fig. 1) are being stored in managed boreal forests on a regular basis as a result of practical forest operations. To our knowledge, this kind of study – that focuses on piles formed in “everyday” forestry – has not been conducted before with piles formed by modern harvesters.
Tomicus beetles used the roundwood piles extensively, as evident by the number of their emergence holes. Of the two species, T. piniperda was responsible for nearly all emergence holes, whereas the occurrence of T. minor was negligible. This is in accordance with previous studies about how the two species differ in relation to the types of material used for breeding (Juutinen 1978; Långström 1984; Borkowski and Skrzecz 2016). Differences were also found in how T. piniperda used the logs: emergence holes were most abundant in sections of the logs with a thick bark, and a larger diameter further increased the average number of holes (Table 4), which is also in agreement with past studies on the ecology of the species (Borkowski and Skrzecz 2016; Juutinen 1978). The proportion of damaged bark clearly made the log less usable for the beetles (Table 3, Fig. 5). Modern harvester heads commonly debark the log, and therefore, it is valid to ask whether the damage risk by the roundwood piles can be decreased by ensuring a higher percentage of harvester-damaged bark, especially in logs placed on the top-most layer of the pile. Based on our results, a 50% decrease in the level of intact bark seemed to decrease the number of emerged beetles by up to 100% (Table 3, Fig. 5).
In our study, the highest numbers of beetle exit holes were over 1000 per m2, and the pile-wise averages varied between 80 and 592 exit holes per m2 (Table 4). In an earlier Finnish study Juutinen (1978) reported maximum densities of 200–300 holes per m2, whilst the pile-wise averages were between 10 and 20 holes per m2. Annila and Petäistö (1978) reported a maximum of approximately 500 exit holes per m2 in windthrown Scots pines. In Sweden, Långström (1984) recorded infestation levels as high as several thousand beetles per tree from windthrown Scots pines and estimated that offspring production was nearly 1000 beetles per m2. Yet, despite the partly damaged bark, the numbers of emerged insects in our study were considerably higher than those in the study by Juutinen (1978), which could be explained by the more southernly location of our study sites. The location of our study sites was something we were not able to affect: the companies were responsible for deciding which piles they were able to spare for research purposes. Therefore, the potential effect of the geographical location on the results cannot be fully answered by our study. However, the modelling made it clear that differences among the piles were more significant than differences between the survey years (Table 3, Table 4), suggesting that local conditions do play a role in determining the local damage risk.
In any case, a considerable number of insects did emerge from the piles, causing shoot damage in the surrounding forests. According to the modelling, the damage was driven predominantly by the volume of the pile and the distance from the pile (Table 5, Figs. 6 and 7). In most cases, the damages ceased when distances to the piles exceeded 40–60 m (Figs. 6 and 7). The structure of the surrounding forest had no significant effect on the level of shoot damage: all variables describing site type, or the density and structure of the forest were omitted in the variable selection phase due to being insignificant. This is not surprising as pine shoot beetles are highly common in Finland, and they can be expected to be present in any forest dominated by Scots pine. Because of this, the structure of the surrounding forest was not assumed to be the main driver of the beetle numbers in the first place. The measurements were important for transforming the plot-level shoot damage into estimates of damage towards a single tree – the measure of potential growth losses. This is needed for assessing the regulations of the current forests legislation for the Tomicus beetles.
To what extent the recorded shoot losses were harmful for the tree growth depends on what is seen as a considerable damage in the first place. Growth losses for young pines have been reported after a loss of 15–30 shoots (Andersson 1974; Nilsson 1974), but for more mature trees, the commonly stated values seem to be between 100 and 200 lost shoots per tree (Nilsson 1974; Stroink 1982; Komonen 2009; Öhrn et al. 2018). Here, the estimated tree-wise shoot losses were, on average, below the literature-cited thresholds. The tree-wise losses exceeded 100 shoots around six piles, and the maximum tree-wise shoot loss was estimated to be 517 fallen shoots per tree (Pile 110). The plots where the highest numbers of fallen shoots were counted were located closer to the piles (Tables 5 and 6). However, the stand surrounding some of the piles with the highest shoot damage had also high amounts of logging residues (which contained galleries of T. minor). It should be noted here that the numbers of fallen shoots that we counted must be treated as underestimates: it has been shown in field studies that shoots damaged by the beetles will fall off also during winter, and that this may double the total number of shoots fallen due to Tomicus damage (Borkowski 2007). Therefore, we must assume that we only counted a portion of the shoots that were damaged. Increasing the number of the counted fallen shoots by e.g. 20% would keep our results as they were in relation to the limit of 100 fallen shoots per tree: the limit would not be exceeded in any other than the six piles where the number of fallen shoots was the highest. For the smallest piles (<50 m3), the number of fallen shoots we counted should be increased by 150% before the limit of 100 fallen shoots would exceed.
Previous studies have estimated that growth losses can last from 1 to up to 10 years. Via simulated pruning, Ericsson et al. (1985) found minor losses in radial growth that were visible 2 years after the study; the maximum number of cut shoots was 200 from a single tree. In a 28-year-old pine forest, i.e., younger than our forests, Stroink (1982) found an 11% reduction in basal area growth after 170 shoots had been lost, but the effect lasted only for 1 year. The most severe effects have been reported when the leading shoot has been attacked: Andersson (1973) reported decreased height growth for up to 18 years. When it comes to volume, loss estimates of up to 8–10 years have been reported in cases where the wood is stored in the same area on a more permanent basis (Långström and Hellqvist 1991). Here, we cannot assess the duration of the potential growth losses.
In operational forestry, the storage of roundwood in piles along forest roads is temporary, meaning that the same resource capitalised by the beetles in one summer will not be there during the next, which limits the local damage, provided that there will be no further roundwood piles near the same stand in the following years. Annila and Heikkilä (1991) studied pine shoot beetle densities and shoot damage across Finland after the thinning of young Scots pine stands and found no notable negative effect on tree growth. In their study on pine shoot beetle abundance and damage after storm, Komonen et al. (2009) reached the same conclusion as the trees felled by the storm in one year were unusable for the beetles in the next year. This, together with our results, suggests that the damage potential of this species does not rise significantly if the extra resources for breeding (such as the roundwood piles) are only temporary. However, pine shoot beetles can cause mortality to other species of pine elsewhere (Ye et al. 2003), and along with many other bark beetles, they are predicted to benefit from a changing climate as warmer summers facilitate the development of larvae into adults and increase the likelihood of a sister brood: adults breeding twice during a single summer (Saarenmaa 1985). Therefore, the role of this species as a damage-causing agent for Scots pine may change in the future, but currently, in the northern conditions it can be viewed as a notable pest agent only around long-term wood storage sites.
In conclusion, based on our findings the present legislation on the storing of roundwood piles of pine seems reasonable as piles smaller than 50 m3 did not cause notable damage; the legislation only mandates the removal of piles with a volume higher than 20 m3. It should be noted that the legislation (and the same limits for the amount of wood in m3) is the same for piled roundwood as well as for trees damaged by snow or wind, and it does not concern dead wood – only wood cut or fallen during the previous autumn–spring period. Therefore, in the event of a storm, it seems reasonable to assume that the fallen pines do not increase damage risks any more than stored roundwood piles, pending that the forest does not become too sparse due to the storm, forcing all the emerging beetles to attack the single remaining trees. Also, our study piles were surrounded by mature pine forests. Had the recorded level of damage occurred in younger forests, growth losses would have been inevitable. Similarly, if a forest surrounding a large pile of roundwood pine is sparse, or if it becomes sparse after a storm, the risk of damage would naturally increase as the beetles emerging from the trunks will attack the nearby trees – the less trees there are, the higher the density of beetle attacks per single tree. Therefore, the question of where to store large piles of roundwood pine is a topical one when it comes to the damage risk: next to a young pine stand or a spruce or birch stand. Equally, the possibility of causing more damages to the bark of the logs by the harvester heads should be explored as a reasonable and clearly as an effective method to decrease the number of beetles breeding in the piles. Nevertheless, the functionality of the forest damage legislation against the background of a changing climate calls for continuous surveillance of the changing conditions with respect to the damaging agents. This study provided practical information for the case of Tomicus beetles and piles of roundwood Scots pine.
Author contributions
HV, AP and SN formulated the field methods and HV and AP collected the 2017 data. TY and MM led the research and field campaign for 2020, together with KH and LA. LA digitized the 2020 field data. MM integrated the 2017 and 2020 datasets, wrote the first draft of the manuscript and conducted the analysis. After that, all authors contributed to the final manuscript.
Acknowledgements
We would like to thank Metsähallitus Metsätalous Oy (the company managing Finnish state-owned forests), Stora Enso Oy, Metsä Group Oyj, Tornator Oyj and UPM-Kymmene Oyj for providing the roundwood piles for the study and the Finnish Forest Centre for the permission to extend the storage time of the study piles. The study was funded by the Ministry of Agriculture and Forestry. We also thank Tapio Ylimartimo, Juha Metros, Martti Vuorinen, Ari Rajala and Raimo Jaatinen for assisting with field work.
Data accessibility statement
The data collected in this study are available from the corresponding author upon reasonable request.
References
Andersson SO (1973) Increment losses after thinning caused by Blastophagus piniperda. Sveriges skogsvdrdsfdrbunds tidskrift 71: 359–379.
Annila E, Heikkilä R (1991) Breeding efficiency in Tomicus piniperda and shoot damage after late autumn thinning of young Pinus sylvestris stands. Scand J Forest Res 6: 197–207. https://doi.org/10.1080/02827589109382661.
Annila E, Petäistö RL (1978) Insect attack on windthrown trees after the December 1975 storm in western Finland. Commun Inst For Fenn 94: 1–24. http://urn.fi/URN:NBN:fi-metla-201207171125.
Borkowski A (2007) Spatial distribution of fallen shoots of Scots pine pruned by pine shoot beetles (Tomicus spp.), and evaluation of methods of shoot collection in central Poland. J For Res 12: 358–364. https://doi.org/10.1007/s10310-007-0029-5.
Borkowski A, Skrzecz I (2016) Ecological segregation of bark beetle (Coleoptera, Curculionidae, Scolytinae) infested in Scots pine. Ecol Res 31: 135–144. https://doi.org/10.1007/s11284-015-1322-y.
Cajander AK (1949) Forest types and their significance. Acta For Fenn 56. https://doi.org/10.14214/aff.7396.
Ericsson A, Hellqvist C, Långström B, Larsson S, Tenow O (1985) Effects on growth of simulated and induced shoot pruning by Tomicus piniperda as related to carbohydrate and nitrogen dynamics in Scots pine. J Appl Ecol 22: 105–124. https://doi.org/10.2307/2403331.
Finlex (2013) Laki metsätuhojen torjunnasta 1087/2013. [The Law on Prevention of Forest Damage]. https://www.finlex.fi/fi/laki/alkup/2013/20131087.
Juutinen P (1978) Pulpwood stacks as breeding sites for pine shoot beetle (Tomicus piniperda L.) in Northern Finland. Folia For 335. http://urn.fi/URN:ISBN:951-40-0315-2.
Kniivilä M, Hantula J, Hotanen J-P, Hynynen J, Hänninen H, Korhonen KT, Leppänen J, Melin M, Mutanen A, Määttä K, Siitonen J, Viiri H, Viitala E-J, Viitanen J (2020) Metsälain ja metsätuholain muutosten arviointi. [Assessing the amendments of the finnish forest legislation and its damage prevention act]. Luonnonvara- ja biotalouden tutkimus 3/2020. http://urn.fi/URN:ISBN:978-952-326-897-5.
Komonen A, Laatikainen A, Similä M, Martikainen P (2009) Ytimennävertäjien kasvainsyönti trombin kaataman suojelumännikön ympäristössä Höytiäisen saaressa Pohjois-Karjalassa. [Shoot damages caused by the pine shoot beetle in a conservation area after storm damage] Metsätieteen aikakauskirja 2/2009: 127–134. https://doi.org/10.14214/ma.5761.
Långström B (1984) Windthrown Scots pines as brood material for Tomicus piniperda and T. minor. Silva Fenn 18: 187–198. https://doi.org/10.14214/sf.a15392.
Långstrom B, Hellqvist C (1991) Shoot damage and growth losses following three years of Tomicus-attacks in Scots pine stands close to a timber storage site. Silva Fenn 25: 133–145. https://doi.org/10.14214/sf.a15604.
Metsäteho (2003) Kuitupuun pinomittaus. ISBN 951-673-184-8. http://www.metsateho.fi/wp-content/uploads/2015/02/Kuitupuun_pinomittaus_ohje_uusi.pdf.
Nilsson S (1974) Increment losses caused by Blastophagus piniperda on Scots pine. Royal College of Forestry, Garpenberg, Research Notes 78.
Öhrn P, Björklund N, Långström B (2018) Occurrence, performance and shoot damage of Tomicus piniperda in pine stands in southern Sweden after storm-felling. J Appl Entomol 142: 854–862. https://doi.org/10.1111/jen.12533.
Pinherio JC, Bates DM (2004) Mixed-effects models in S and S-PLUS. Springer, New York, NY, Statistics and Computing Series. https://doi.org/10.1007/b98882.
Pinheiro J, Bates D, DebRoy S, Sarkar D, R Core Team (2020) nlme: linear and nonlinear mixed effects models. R package version 3.1-148. https://CRAN.R-project.org/package=nlme.
Saarenmaa H (1985) The role of temperature in the population dynamics of Tomicus piniperda (L.) (Col. Scolytidae) in northern conditions. J Appl Entomol 99: 224–236. https://doi.org/10.1111/j.1439-0418.1985.tb01982.x.
Seidl R, Rammer W (2017) Climate change amplifies the interactions between wind and bark beetle disturbances in forest landscapes. Landscape Ecol 32: 1485–1498. https://doi.org/10.1007/s10980-016-0396-4.
Silva X (2011) Studies on fungi associated with Tomicus piniperda L. in Finland. Master’s Thesis, University of Helsinki. http://urn.fi/URN:NBN:fi:hulib-201507211840.
Solheim H, Långström B (1991) Blue-stain fungi associated with Tomicus piniperda in Sweden and preliminary observations on their pathogenicity. Ann Sci Forest 48: 149–156. https://doi.org/10.1051/forest:19910203.
Stroink HJ (1982) Bestandesschäden durch Ernährungsfrass und Brutbefall des grossen Waldgärtners (Blastophagus piniperda L.) an Kiefer (Pinus sylvestris L.) [Stand level damage on pine caused by shoot feeding and brood infestation of the pine shoot beetle] Thesis, Georg-August-Universität, Göttingen.
Wickham H (2016) ggplot2: elegant graphics for data analysis. Springer, Cham. https://doi.org/10.1007/978-3-319-24277-4.
Ye H, Haack R, Lu J (2003) Tomicus piniperda (Scolytidae): a serious pest of Yunnan pine in southwestern China. Proceedings of the 2002 U.S. Department of Agriculture Interagency Research Forum GTR-NE-300. https://www.nrs.fs.fed.us/pubs/gtr/gtr_ne300/gtr_ne300_105.pdf.
Total of 25 references.
