Francesco Chianucci  ,
Luca Salvati,
Tessa Giannini,
Ugo Chiavetta,
Piermaria Corona,
Andrea Cutini
,
Luca Salvati,
Tessa Giannini,
Ugo Chiavetta,
Piermaria Corona,
Andrea Cutini
Long-term response to thinning in a beech (Fagus sylvatica L.) coppice stand under conversion to high forest in Central Italy
Chianucci F., Salvati L., Giannini T., Chiavetta U., Corona P., Cutini A. (2016). Long-term response to thinning in a beech (Fagus sylvatica L.) coppice stand under conversion to high forest in Central Italy. Silva Fennica vol. 50 no. 3 article id 1549. https://doi.org/10.14214/sf.1549
Highlights
- Canopy recovery after medium-heavy thinning reveals the prompt response of beech to intensive thinning cycles
- Active management practices accelerate the transition from coppice to high forest.
Abstract
European beech (Fagus sylvatica L.) forests have a long history of coppicing, but the majority of formerly managed coppices are currently under conversion to high forest. The long time required to achieve conversion requires a long-term perspective to fully understand the implication of the applied conversion practices. In this study, we showed results from a long-term (1992–2014) case-study comparing two management options (natural evolution and periodic thinning) in a beech coppice in conversion to high forest. Leaf area index, litter production, radiation transmittance and growth efficiency taken as relevant stand descriptors, were estimated using both direct and indirect optical methods. Overall, results indicated that beech coppice showed positive and prompt responses to active conversion practices based on periodic medium-heavy thinning. A growth efficiency index showed that tree growth increased as the cutting intensity increased. Results from the case study supported the effectiveness of active conversion management from an economic (timber harvesting) and ecological (higher growth efficiency) point of view.
Keywords
continuous cover forestry;
thinning;
Leaf Area Index;
coppice management;
Plant Canopy Analyzer
-
Chianucci,
Consiglio per la Ricerca in Agricoltura e l’Analisi dell’Economia Agraria – Forestry Research Centre, viale Santa Margherita 80, 52100 Arezzo, Italy
 http://orcid.org/0000-0002-5688-2060
E-mail
fchianucci@gmail.com
http://orcid.org/0000-0002-5688-2060
E-mail
fchianucci@gmail.com
- Salvati, Consiglio per la Ricerca in Agricoltura e l’Analisi dell’Economia Agraria – Research Centre for the Soil-Plant System, via della Navicella 2–4, 00184 Roma, Italy E-mail bayes00@yahoo.it
- Giannini, Consiglio per la Ricerca in Agricoltura e l’Analisi dell’Economia Agraria – Forestry Research Centre, viale Santa Margherita 80, 52100 Arezzo, Italy E-mail tessa.giannini@entecra.it
- Chiavetta, Consiglio per la Ricerca in Agricoltura e l’Analisi dell’Economia Agraria – Forestry Research Centre, viale Santa Margherita 80, 52100 Arezzo, Italy E-mail ugo.chiavetta@entecra.it
- Corona, Consiglio per la Ricerca in Agricoltura e l’Analisi dell’Economia Agraria – Forestry Research Centre, viale Santa Margherita 80, 52100 Arezzo, Italy E-mail piermaria.corona@unitus.it
- Cutini, Consiglio per la Ricerca in Agricoltura e l’Analisi dell’Economia Agraria – Forestry Research Centre, viale Santa Margherita 80, 52100 Arezzo, Italy E-mail andrea.cutini@entecra.it
Received 11 January 2016 Accepted 2 May 2016 Published 6 May 2016
Views 114550
Available at https://doi.org/10.14214/sf.1549 | Download PDF

Supplementary Files
1 Introduction
Beech (Fagus sylvatica L.) is an economically and ecologically important tree species in Europe (Bolte et al. 2007), where it covers about 15 million hectares (FOREST EUROPE, UNECE and FAO 2011). Although the species occurs from the plains to about 1800 meters above sea level throughout its European range, its distribution prevails in mountain areas of Central and Southern Europe. The cultivation of beech forest has represented a fundamental resource for people living in mountain areas where human practices have significantly modified the distribution, composition and structure of beech forests (Brunet et al. 2010; Nocentini 2009; Scarascia-Mugnozza et al. 2000). In particular, most beech forests in Southern Europe had a long history of coppicing, a situation very common in Italian beech forests. Economic and social changes in the last decades have supported widespread conversion of many coppices into high forests, a situation very common in many European countries (Cañellas et al. 2004; Chianucci et al. 2015b; Cutini et al. 2013; Di Matteo et al. 2010). At present time, stored coppices (i.e., stands left growing beyond the customary coppice rotation) represent 89% of the whole coppice area in Italy, and managed coppices in transition to high forest occupy about 150 000 ha (NFI 2005).
Currently, the management situation of beech forests in Italy is quite diversified. While high beech forests prevail in Southern Italy and in the Alps, most beech stands originated from coppicing are mainly located in Northern and Central Apennines (61%). On one hand, conversion has usually been carried out by gradually thinning the stand; this option has commonly been adopted in public areas (Ciancio et al. 2006). On the other hand, conversion to high forest has not been so widespread in private forests and there are large areas where coppices have been abandoned (Nocentini 2009). The co-existence of large areas (roughly 500 000 ha) either abandoned or under active conversion raises several questions concerning present and future management perspectives for beech coppice woods.
Whatever the management, the ecological implications of coppice conversion is still poorly documented. As the conversion in mountain area is a long process, a long-term perspective is required to define appropriate management options to fulfil conversion targets. Furthermore, analysing the effects of different management options in an objective and reproducible way is usually challenging, because different methodologies and sampling strategies have been proposed, severely hampering comparisons among studies (Franklin et al. 2002). This calls for an evaluation of the different management options using objective methods and techniques. This study is a part of a wider long-term project focusing on designing different silvicultural options for managing pure beech stands (Cutini et al. 2015, but see also the LIFE14 ENV/IT/000514 project: www.futureforcoppices.eu). Specifically, in the present study we investigated the effects of two silvicultural alternatives (natural evolution and active conversion) in a former coppice stand, which was monitored during a 23 years period. The analysis included canopy attributes and growth efficiency measured from well-established forestry techniques. Finally, we interpreted the results based on an integrated approach to outline guidelines for managing beech forests originating from coppice.
2 Material and methods
2.1 Study area and experimental design
The study was conducted in a mountainous area of the Northern Apennines, Italy (Alpe di Catenaia; 43°49´N, 11°49´E). The experimental trial consists of two permanent pure beech plots established in a previous study (Amorini et al. 2010) in 1972, including the natural evolution pattern (unthinned control) and the traditional conversion system (periodic thinning). The plots were 1 ha in size each and were chosen within a uniformly managed coppice stand aged 27, i.e. at a time around the traditional rotation. Four inventories were carried out in both plots since 1972 at each decade up to 2012 (Table 1). The original experimental design included replicates for assessing growth dynamics and mortality in the different management options (see Amorini et al. 2010). However, to get a compromise between long-term monitoring of the effect of the silvicultural alternatives on beech and the annual survey of a number of canopy attributes (see 2.2), we were forced to not consider replicates in the current experiment.
| Table 1. Comparison of stand characteristics measured in formerly managed beech coppices at the end of the rotation period (27 years) in 1972. The two plots have been started of being converting since 1972 to high forest following two management options: natural evolution (unthinned control) and periodic thinning. For the thinned plot, data are reported, either before (bt) or after (at) each thinning. The reduction in stools and basal area (%) is also reported (brackets); dg: quadratic mean diameter; ddom: top diameter; hdom: dominant height. | ||||||
| ID | Inventory | Stools | Basal area | dg | ddom | hdom |
| Control | 1972 | 7518 | 27.4 | 6.8 | 18.6 | 11.2 |
| 1982 | 6846 | 38.2 | 8.4 | 23.0 | 15.7 | |
| 1987 | 3920 | 40.9 | 11.5 | 29.0 | 20.1 | |
| 2002 | 2580 | 45.3 | 15.0 | 33.0 | 25.1 | |
| 2012 | 2046 | 48.15 | 17.3 | 35.2 | 26.7 | |
| Thinned | 1972 bt | 7407 | 27.5 | 6.8 | 18.6 | 11.2 |
| 1972 at | 2222 (–70%) | 17.3 (–37%) | 9.6 | 15.9 | 10.7 | |
| 1982 | 2182 | 29.4 | 12.8 | 22.1 | 15.7 | |
| 1987 bt | 2076 | 35.2 | 14.1 | 24.9 | 17.6 | |
| 1987 at | 706 (–66%) | 19.7 (–44%) | 18.8 | 24.8 | 17.6 | |
| 2002 bt | 702 | 33.5 | 24.6 | 33.6 | 24.2 | |
| 2002 at | 414 (–41%) | 23.8 (–29%) | 27.1 | 33.5 | 24.2 | |
| 2012 | 412 | 29.8 | 30.4 | 37.5 | 25.6 | |
According to the classical French tradition (Hubert 1983), active conversion practices consist in gradually reducing the number of shoots by means of periodic low thinning, with the first thinning applied close to the end of the coppice rotation. The interval between two consecutive thinning procedures was 15–20 years, with a temporary opening of the canopy cover, to enable seed regeneration under gradually decreasing cover. Regeneration cutting conclude this conversion system, which can last up to 150 years, (Hahn and Fanta 2001). In our stands, three medium-heavy thinnings were done in 1972, 1987 and September 2002 in the thinned plot. Based on definition of optimal target stand density (Ciancio et al. 2006), the adopted thinning reduced the number of stools and the basal area to values ranging from 41% to 70% and from 29% to 44%, respectively (see Table 1).
2.2 Canopy properties and growth efficiency estimates
The following canopy attributes have been estimated in the study plots using various methods: litter production and its partitioning, leaf area index and canopy transmittance ; we used pseudo-replicates to calculate these attributes at plot level. Growth efficiency was also calculated in each plot.
Litter production was estimated using litter traps following the protocol by Chianucci and Cutini (2013). Nine 0.25 m2 traps were placed on a grid within each plot at a height of 1 m. Litter was collected during 1992–2014; the collection was conducted every year at intervals of 10–15 days during October–January. Litter was grouped into leaves, woody and reproductive parts; and dried at 85°±2 for 24 hours in the laboratory using forced air stoves. The necromass values (Mg ha−1) of each litter component were determined using digital scale with ±1 g precision.
Leaf area index estimates were performed every year during 1999–2014 in July–August under diffuse and uniform sky conditions, using a Plant Canopy Analyzer LAI-2000 PCA (Li-Cor Inc., Lincoln, USA), equipped with a 90° azimuth view cap. The optics of PCA consist of a fisheye lens (148° field ofview) divided into five concentric rings (with central zenith angle of 7°, 23°, 38°, 53°, and 68°, respectively). Each ring simultaneously integrates incoming radiation in the 320–490 nm wavelength range (blue light) over the view cap azimuth range selected. One above-canopy reference measurement for each plot was recorded in clearings near each study area. Nine below-canopy measurements were recorded within each plot at the same grid points used for litter collection. Leaf area estimates were calculated from a radiative formula. We omitted the largest zenith angle (LAI4) to reduce woody contribution on below-canopy measurements
Canopy direct transmittance was measured every year during 1992–2014 in July–August at noon under clear sky conditions with an AccuPAR ceptometer (Decagon Devices, Pullman, USA). The instrument consists of an integrated microprocessor-driven data logger and probe. The probe contains 80 independent sensors, spaced 1 cm apart. The photosensors measure PAR (Photosynthetically Active Radiation) in the 400–700 nm waveband (µmol m−2s−1). One above-canopy measurement for each plot was recorded in clearings near each study area. Nine below-canopy measurements were done within each plot at the same grid points used for litter collection. Four measurements per sampling point were recorded, for a total of 36 measurements (2880 PAR readings) per plot. We calculated the transmittance (Trasm) as the percentage between the annual averaged below light measurement to the above radiation measured in each plot.
Growth efficiency (ω) was calculated as
where ∆G is the stand basal area increment (calculated between the inventories 1987–2002 and 2003–2014) and LAI4 is the mean stand leaf area index of each plot (estimated using LAI-2000 PCA over 1999–2002 and 2003–2014).
2.3 Statistical analyses
The impact of the two management options on canopy attributes was evaluated using linear mixed effects (LME) model, with year as a random effect and silvicultural option as a fixed effect. Statistical analyses were performed in R version 3.03 (R Development Core Team 2013) with the ‘lmerTest’ library uploaded.
3 Results
Table 2 illustrates average values obtained from the canopy variables in the natural evolution and thinned plots. With regard to litter production, leaves represented the major litterfall component, comprising 77% and 72% of total litter in thinned and unthinned plots, respectively. Woody elements formed the second most important component, 13% and 16% of litterfall of thinned and unthinned plots, respectively. Reproductive parts contributed approximately 9–10% of total litter in both plots. Non-classified elements were 1–2% of the total litter.
| Table 2. (Top): Statistics on annual litter production and its partitioning (measured during1992–2014), leaf area index (LAI4; 1999–2014) and canopy transmittance (Trasm; 1999–2014) in control and thinned plots. The number of sampling points in both plots was 9 for litterfall and LAI4 measurements and 36 for canopy transmittance measurements. (Bottom): Growth efficiency estimates obtained after comparison of inventories (1987–2002; 2003–2014) at stand (Estand) and individual tree level (Etree). ∆G: basal area increment between the considered inventories. | |||||||||
| Stand ID | Litter components (Mg ha–1) | LAI4 (m2 m–2) | Trasm. % | ||||||
| Leaves | Woody | Nuts | Husks | Other | Total | ||||
| Control | Average | 3.785 | 0.858 | 0.135 | 0.331 | 0.127 | 5.237 | 6.1 | 1.2 |
| SE | 0.089 | 0.081 | 0.041 | 0.079 | 0.031 | 0.159 | 0.1 | 0.2 | |
| Min | 2.760 | 0.278 | <0.001 | 0.010 | <0.001 | 3.757 | 4.5 | 0.3 | |
| Max | 4.403 | 1.640 | 0.888 | 1.672 | 0.481 | 6.631 | 7.0 | 3.0 | |
| CV | 11.3 | 45.1 | 144.5 | 114.6 | 116.4 | 14.5 | 10.9 | 59.5 | |
| Thinned | Average | 3.341 | 0.544 | 0.115 | 0.294 | 0.049 | 4.346 | 5.0 | 4.0 |
| SE | 0.132 | 0.086 | 0.044 | 0.085 | 0.019 | 0.189 | 0.2 | 0.9 | |
| Min | 1.889 | 0.097 | <0.001 | <0.001 | <0.001 | 2.619 | 2.3 | 0.4 | |
| Max | 4.709 | 1.671 | 0.982 | 1.780 | 0.321 | 6.311 | 6.7 | 16.7 | |
| CV | 18.9 | 76.0 | 183.3 | 137.9 | 186.8 | 20.8 | 21.1 | 108.5 | |
| Stand ID | 1987–2002 | 2003–2014 | |||||||
| ∆G | LAI4 | Estand | Etree | ∆G | LAI4 | Estand | Etree | ||
| Control | 4.4 | 5.9 | 0.74 | 0.0003 | 2.9 | 6.1 | 0.46 | 0.0002 | |
| Thinned | 13.8 | 5.6 | 2.47 | 0.0035 | 6.0 | 4.8 | 1.26 | 0.0030 | |
The last thinning (2001) reduced total litter from 5.762 Mg ha−1 to 5.056 Mg ha−1 and leaf litter from 4.709 Mg ha−1 to 2.767 Mg ha−1 (Fig. 1). Conversely, woody elements increased from 0.546 Mg ha−1 in 2001 to 1.671 Mg ha−1 in 2002, most likely because of rapid stress response after thinning (Fig. 1). Leaf, woody and total litter was significantly lower in the thinned stand compared with the unthinned control (LME, p < 0.05). Conversely, reproductive parts were not significantly different in the two plots.
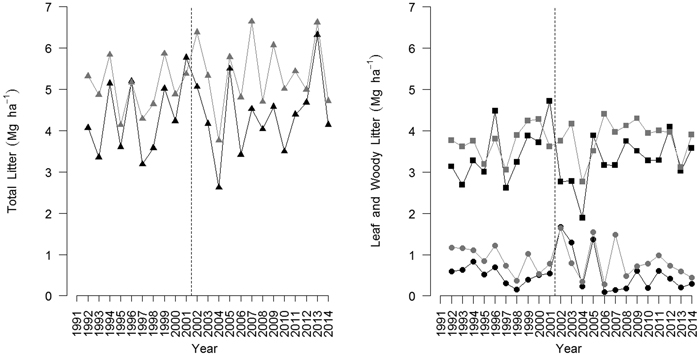
Fig. 1. Time pattern of total litter (triangles), leaf (squares) and woody (circles) litter production in the beech coppices under conversion to high forests following two management options: periodic thinning (black) and unthinned control (grey). The vertical dashed line indicates the last thinning undertaken in the thinned stand. View larger in new window/tab.
Consistently with litter removal, thinning reduced LAI4 from 5.66 m2 m−2 in 2002 to 4.13 m2 m−2 in the subsequent year (Fig. 2). LAI4 in the thinned stand was significantly lower than that in the unthinned stand (LME, p < 0.05), even though the differences appeared to plateau rapidly over time.
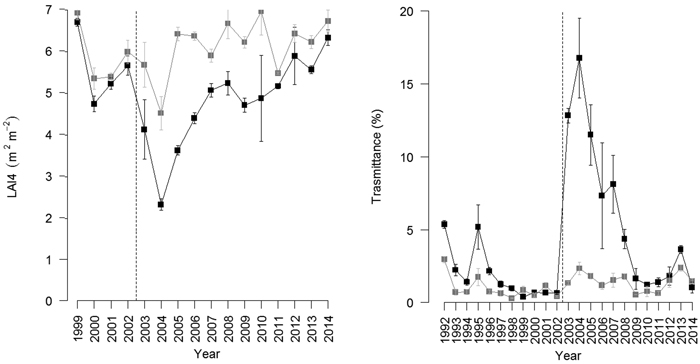
Fig. 2. (Left): Time pattern of leaf area index calculated from LAI-2000 PCA (LAI4, excluding the outer ring) in the beech coppices under conversion to high forests following two management options: periodic thinning (black) and unthinned control (grey). The vertical dashed line indicates the last thinning undertaken in the thinned stand. (Right): Time pattern of canopy transmittance in thinned (black) and unthinned (grey) stands from 1992–2014. The vertical dashed line indicates the last thinning undertaken in the thinned stand. View larger in new window/tab.
An opposite trend was observed for radiation transmittance (Trasm). The last thinning increased canopy transmittance from 0.66% in 2002 to 12.81% in the subsequent year (Fig. 2). Canopy transmittance in the thinned stand was not significantly different from that in the unthinned stand before thinning, but was significantly higher in the period after thinning (LME, p < 0.05), although the differences appeared to plateau quickly with time.
Growth efficiency (ω) estimated at two periods indicated that stand growth rate and efficiency increased with thinning, resulting in larger individual tree growth in the retained trees (Table 2), as also suggested by the quadratic mean diameter obtained in the thinned plot (Table 1).
4 Discussion
In this study we compared the implication of two alternative management options in beech coppice over more than two decades. All statistical tests indicated that the surveyed variables are influenced by the silvicultural treatments in the long-term, supporting the reliability of the monitored variables to assess the beech response to the silvicultural interventions (Salvati and Zitti 2009). In addition, the sampling protocol was comparable with that adopted in previous studies (e.g. Genet et al. 2010; Lebret et al. 2001; Leuschner et al. 2006; Meier and Leuschner 2008), allowing thorough evaluation of the different conversion options in beech forests. It should, however, be stressed that our study was limited to a cause-study level. Even if we judged the canopy properties estimates obtained herein as reliable average values for beech, on account of the relatively large time series obtained, the lack of replicates implies that generalisation of the outcome should be carefully evaluated.
The analyses presented here indicated the high productivity of the stands investigated, as outlined by the average values of litterfall and LAI4, which are comparable with previous reports (Cutini 2002; Lebourgeois et al. 2005; Matteucci et al. 1999; Meier and Leuschner 2008; Chianucci et al. 2015a; Chianucci 2016). This result is important because medium-high stand productivity is a pre-requisite for achieving successful conversion management.
Leaves represented the main litter component in both plots. Accordingly, leaf litter and its related attributes such as LAI4 and Trasm were significantly affected by the two management options, even though the differences between the two plots were the highest in the years immediately after thinning and appeared to level out with time, indicating the prompt response of beech to intensive thinning cycles, as observed for other broadleaves (Rodriguez-Calcerrada et al. 2011). The growth efficiency indexes further confirmed that thinning increased the growth performance, most likely due to the reduction in competition for light and the subsequent recovery of available space (Cescatti and Piutti 1998; Van der Maaten 2013).
In summary, our study demonstrated the effectiveness of active conversion practices for the management of beech coppices. Compared with natural evolution, the active management is profitable due to frequent harvest (Ciancio et al. 2006; Amorini et al. 2010) while simultaneously yielding more growth-efficient trees; this effect, in turn contributes to accelerate the transition of thinned plot to more adult stand conditions, as larger growing space allows larger crown enlargement of retained trees, encouraging larger seed production and light at the forest floor to favour seed regeneration (Chianucci et al. 2014).
Acknowledgments
Francesco Chianucci and Luca Salvati designed the experiment, coordinated the research and wrote the paper; other co-authors provide conceptual, field data collection and logistic support. We are grateful to the late Germano Gambi, Emilio Amorini and Gianfranco Fabbio for setting up the experimental sites. We are grateful to collaborators and technicians of Forestry Research Centre who contributed to data collection. The authors also wish to thank the anonymous reviewers for their helpful suggestions.
Financial support was provided by the LIFE Project FutureForCoppiceS (LIFE14 ENV/IT/000514), www.futureforcoppices.eu. Francesco Chianucci was also funded by the research project “ALForLab” (PON03PE_00024_1) co-funded by the (Italian) National Operational Programme for Research and Competitiveness (PON R&C) 2007–2013, through the European Regional Development Fund (ERDF) and national resource (Revolving Fund - Cohesion Action Plan (CAP) MIUR).
References
Amorini E., Fabbio G., Bertini G. (2010). Dinamica del ceduo oltre turno e avviamento ad alto fusto dei cedui di faggio. Risultati del protocollo “Germano Gambi” sull’Alpe di Catenaia (Arezzo). Annals of Silvicultural Research 36: 151–172. [In Italian].
Bolte A., Czajkowski T., Kompa T. (2007). The north-eastern distribution range of European beech – a review. Forestry 80(4): 413–429. http://dx.doi.org/10.1093/forestry/cpm028.
Brunet J., Fritz Ö., Richnau G. (2010). Biodiversity in European beech forests – a review with recommendations for sustainable forest management. Ecological Bulletins 53:77–94.
Cañellas I., Del Rìo M., Roig S., Montero G. (2004). Growth response to thinning in Quercus pyrenaica Willd. coppice stands in Spanish central mountain. Annals of Forest Sciences 61(3): 243–250. http://dx.doi.org/10.1051/forest:2004017.
Cescatti A., Piutti E. (1998). Silvicultural alternatives, competition regime and sensitivity to climate in a European beech forest. Forest Ecology and Management 102(2–3) : 213–223. http://dx.doi.org/10.1016/S0378-1127(97)00163-1.
Chianucci F. (2016). A note on estimating canopy cover from digital cover and hemispherical photography. Silva Fennica 50(1) article 1518. http://dx.doi.org/10.14214/sf.1518.
Chianucci F., Cutini A. (2013). Estimation of canopy properties in deciduous forests with digital hemispherical and cover photography. Agricultural and Forest Meteorology 168: 130–139. http://dx.doi.org/10.1016/j.agrformet.2012.09.002.
Chianucci F., Puletti N., Venturi E., Cutini A., Chiavetta U. (2014). Photographic assessment of overstory and understory leaf area index in beech forests under different management regimes in Central Italy. Forestry Studies 61(1): 27–34. http://dx.doi.org/10.2478/fsmu-2014-0008.
Chianucci F., Macfarlane C., Pisek J., Cutini A., Casa R. (2015a). Estimation of foliage clumping from the LAI-2000 Plant Canopy Analyzer: effect of view caps. Trees 29(2): 355–366. http://dx.doi.org/10.1007/s00468-014-1115-x.
Chianucci F., Mattioli L., Amorini E., Giannini T., Marcon A., Chirichella R., Apollonio M., Cutini A. (2015b). Early and long-term impacts of browsing by roe deer in oak coppiced woods along a gradient of population density. Annals of Silvicultural Research 39(1): 32–36. http://dx.doi.org/10.12899/asr-945.
Ciancio O., Corona P., Lamonaca A., Portoghesi L., Travaglini D. (2006). Conversion of clearcut beech coppices into high forests with continuous cover: a case study in central Italy. Forest Ecology and Management 224(3): 235–240. http://dx.doi.org/10.1016/j.foreco.2005.12.045.
Cutini A. (2002). Litterfall and leaf area index in the CONECOFOR permanent monitoring plots. Journal of Limnology 61(1): 62–68. http://dx.doi.org/10.4081/jlimnol.2002.s1.62.
Cutini A., Chianucci F., Chirichella R., Donaggio E., Mattioli L., Apollonio M. (2013). Mast seeding in deciduous forests of the northern Apennines (Italy) and its influence on wild boar population dynamics. Annals of Forest Sciences 70(5): 493–502. http://dx.doi.org/10.1007/s13595-013-0282-z.
Cutini A., Chianucci F., Giannini T., Manetti M.C., Salvati L. (2015). Is anticipated seed cutting an effective option to accelerate transition to high forest in European beech (Fagus sylvatica L.) coppice stands? Annals of Forest Sciences 72(5): 631–640. http://dx.doi.org/10.1007/s13595-015-0476-7.
Di Matteo G., De Angelis P., Brugnoli E., Cherubini P., Scarascia-Mugnozza G. (2010). Tree-rings Δ13C reveals the impact of past forest management on water-use efficiency in a Mediterranean oak coppice in Tuscany (Italy). Annals of Forest Sciences 67(5): 510–518. http://dx.doi.org/10.1051/forest/2010012.
FOREST EUROPE, UNECE and FAO (2011). State of the Europe’s Forests 2011. Status and trends in sustainable forest management in Europe. 341 p.
Franklin J.F., Spies T.A., Pelt R.V., Carey A.B., Thornburgh D.A., Berg D.R., Lindenmayer D.B., Harmon M.E., Keetons W.S., Shaw D.C., Bible K., Chen J. (2002). Disturbances and structural development of natural forest ecosystems with silvicultural implications using Douglas-fir forests as an example. Forest Ecology and Management 155(1–3): 399–423. http://dx.doi.org/10.1016/S0378-1127(01)00575-8.
Genet H., Bréda N., Dufrêne E. (2010). Age-related variation in carbon allocation at tree and stand scales in beech (Fagus sylvatica L.) and sessile oak. (Quercus petraea (Matt.) Liebl.) using a chronosequence approach. Tree Physiology 30(2): 177–192. http://dx.doi.org/10.1093/treephys/tpp105.
Hahn K., Fanta J. (2001). Contemporary beech forest management in Europe. Working report 1. 175 p. http://curis.ku.dk/ws/files/49651691/working_report_1.pdf.
Hubert M., (1983). L’amélioration des taillis par balivage intensif. Institut pour le Development Forester, Paris. 117 p.
Lebourgeois F., Bréda N., Ulrich E., Granier A. (2005). Climate-tree-growth relationships of European beech (Fagus sylvatica L.) in the French Permanent Plot Network (RENECOFOR). Trees 19(4): 385–401. http://dx.doi.org/10.1007/s00468-004-0397-9.
Lebret M., Nys C., Forgeard F, (2001). Litter production in an Atlantic beech (Fagus sylvatica L.) time sequence. Annals of Forest Sciences 58(7): 755–768. http://dx.doi.org/10.1051/forest:2001161.
Leuschner C., Voß S., Foetzki A., Clases Y. (2006). Variation in leaf area index and stand leaf mass of European beech across gradients of soil acidity and precipitation. Plant Ecology 186(2): 247–258. http://dx.doi.org/10.1007/s11258-006-9127-2.
Matteucci G., De Angelis P., Dore S., Masci A., Valentini R., Scarascia-Mugnozza G. (1999). Il bilancio del carbonio delle faggete: dall’albero all’ecosistema. In: Scarascia Mugnozza G. (ed.). Ecologia strutturale e funzionale di faggete italiane, Edagricole, Bologna. p. 133–138.
Meier I.C., Leuschner C. (2008). Leaf size and Leaf Area Index in Fagus sylvatica forests: competing effects of precipitation., temperature and nitrogen availability. Ecosystems 11(5): 655–669. http://dx.doi.org/10.1007/s10021-008-9135-2.
NFI - National Forest Inventory of forests and forest carbon stock (2005). Ministero delle Politiche Agricole, Alimentari e Forestali, Rome, Italy, 653 p. [In Italian].
Nocentini S. (2009). Structure and management of beech (Fagus sylvatica L.) forests in Italy. iForest- Biogeosciences and Forestry 2: 105–113.
Rodríguez-Calcerrada J., Pèrez-Ramos I.M., Ourcival J.M., Limousin J.M., Joffre R., Rambal S. (2011). Is selective thinning an adequate practice for adapting Quercus ilex coppices to climate change? Annals of Forest Sciences 68: 575–585. http://dx.doi.org/10.1007/s13595-011-0050-x.
Salvati L., Zitti M. (2009). The environmental “risky” region: identifying land degradation processes through integration of socio-economic and ecological indicators in a multivariate regionalization model. Environmental Management 44: 888–898. http://dx.doi.org/10.1007/s00267-009-9378-5.
Scarascia-Mugnozza G., Oswald H., Piussi P., Radoglou K. (2000). Forests of the Mediterranean region: gaps in knowledge and research needs. Forest Ecology and Management 132(1): 97–109. http://dx.doi.org/10.1016/S0378-1127(00)00383-2.
Van der Maaten E. (2013). Thinning prolongs growth duration of European beech (Fagus sylvatica L.) across a valley in southwestern Germany. Forest Ecology and Management 306: 135–141. http://dx.doi.org/10.1016/j.foreco.2013.06.030.
Total of 31 references.
