Āris Jansons  ,
Roberts Matisons,
Virgilijus Baliuckas,
Līga Purina,
Oskars Krišāns,
Jānis Jansons,
Imants Baumanis
,
Roberts Matisons,
Virgilijus Baliuckas,
Līga Purina,
Oskars Krišāns,
Jānis Jansons,
Imants Baumanis
Performance variation of lodgepole pine provenances in Latvia
Jansons Ā., Matisons R., Baliuckas V., Purina L., Krišāns O., Jansons J., Baumanis I. (2018). Performance variation of lodgepole pine provenances in Latvia. Silva Fennica vol. 52 no. 5 article id 10014. https://doi.org/10.14214/sf.10014
Highlights
- Performance of 36 provenances of lodgepole pine in 14 trials was studied
- The 29 year survival was ca. 40%
- Provenance and provenance × trial interaction affected dimensions of lodgepole pine
- Provenances from lower latitudes were the most productive.
Abstract
Lodgepole pine (Pinus contorta var. latifolia Engelm.) has been extensively introduced in Scandinavia on less productive sites. Under a changing climate, it also has a high potential in the eastern Baltic region; still, its performance there has scarcely been reported. This study investigated the performance of 36 Canadian provenances in 14 trials in western Latvia. Tree dimensions showed notable provenance and provenance-by-environment variation, implying that local selection by provenance can be applied for improved yield. Southern provenances showed the best height growth, while southwestern (more oceanic) provenances excelled in diameter growth. Most of the quality traits were affected by provenance or provenance-by-environment interaction, yet the variation was lower than for the growth traits.
Keywords
provenance trial;
Pinus contorta var. latifolia;
growth traits;
transfer;
spike knots
-
Jansons,
Latvian State Forest Research Institute ”Silava”, Department of Forest Tree Breeding, Rigas St.t. 111, Salaspils LV-2169, Latvia
E-mail
aris.jansons@silava.lv
- Matisons, Latvian State Forest Research Institute ”Silava”, Department of Forest Tree Breeding, Rigas St.t. 111, Salaspils LV-2169, Latvia E-mail robism@inbox.lv
- Baliuckas, Forest Institute, Lithuanian Centre for Agriculture and Forestry, Department of Forest Tree Genetics and Breeding, Liepu St. 1, Girionys, LT-53101 Kaunas distr., Lithuania E-mail virgilijus.baliuckas@mi.lt
- Purina, Latvian State Forest Research Institute ”Silava”, Department of Forest Tree Breeding, Rigas St.t. 111, Salaspils LV-2169, Latvia E-mail liga.purina@silava.lv
- Krišāns, Latvian State Forest Research Institute ”Silava”, Department of Forest Tree Breeding, Rigas St.t. 111, Salaspils LV-2169, Latvia E-mail oskars.krisans@silava.lv
- Jansons, Latvian State Forest Research Institute ”Silava”, Department of Forest Tree Breeding, Rigas St.t. 111, Salaspils LV-2169, Latvia E-mail janis.jansons.silava@gmail.com
- Baumanis, Latvian State Forest Research Institute ”Silava”, Department of Forest Tree Breeding, Rigas St.t. 111, Salaspils LV-2169, Latvia E-mail imants.baumanis@silava.lv
Received 20 June 2018 Accepted 22 November 2018 Published 3 December 2018
Views 74991
Available at https://doi.org/10.14214/sf.10014 | Download PDF

1 Introduction
Introductions of exotic tree species have been made to maximise yields and the sustainability of stands (Savill 1997). The resistance of species to weather extremes, which are projected to intensify (IPCC 2013), has been identified as a major issue (Schlyter et al. 2006). In Northern Europe, particularly Fennoscandia, lodgepole pine (Pinus contorta var. latifolia Engelm.) is among the few systematically introduced species aiming for improvement of yields, especially in less productive sites, thus meeting the increasing demand for timber (Lindgren 1993; Elfving et al. 2001). The species has been productive, especially at a young age, mostly exceeding local Scots pine (Pinus sylvestris L.) (Larsen and Nielsen 1982; Stephan 1982; Elfving et al. 2001; Kuznetsova et al. 2009), thus aiding for the reduction of rotation period (Elfving et al. 2001), while maintaining comparable wood properties (Sable et al. 2012). At present in Scandinavia, application of lodgepole pine in commercial forestry has been paused due to concerns about potential biotic risks (Despain 2001); still, considering the growing demand for timber, wider use of the species might be resumed (Nilsson et al. 2011).
Tree species with a wide distribution range, including lodgepole pine (Little 1979), have evolutionarily adapted to diverse environments (Fazekas and Yeh 2001). Accordingly, the selection optimally adapted provenances can maximise the performance and sustainability of stands (Ying 1991; Bolte et al. 2009). In Fennoscandia, provenances of lodgepole pine from British Columbia have shown the best performance (Elfving et al. 2001), while provenances from Oregon, Washington and southern parts of British Columbia shown better performance in Germany and Poland (Larsen and Nielsen 1982; Stephan 1982).
In the Baltic States, growth trials of lodgepole pine provenances were established during 1970–1990, yet few results of these trials have been published (Kuznetsova et al. 2009; Jansons et al. 2013b). Considering long-term decisions in forestry, such results are necessary for wider commercial application of the species. The regional specifics in tree growth (Lindner et al. 2010) also imply that local data are essential. The aim of this study was to quantify the variance in the growth and quality traits of lodgepole pine in Latvia for assessment of their potential improvement by selection among provenances.
2 Materials and methods
2.1 Trials and measurements
The study was based on 14 field trials of lodgepole pine established in Latvia (56°34´–57°16´N, 22°12´–24°09´E) during 1979–1994 with one- and two-year-old seedlings to test 36 Canadian provenances (Table 1). The trials had a randomised complete-block design with 4–8 replications and 12–48 tree plots of 3–13 provenances. Most of the provenances were represented in less than four trials (Table 1). The initial spacing of the trees was 2×1 m, yet in the older trials thinning (leaving ca. 40 of trees) was performed at the age 20–22 years. The trials were growing in lowland conditions (elevation < 60 m a.s.l.) on sandy oligotrophic soils and flat topography. The mean annual temperature (±confidence interval) was 6.2 ± 0.3 °C, and the mean monthly temperature ranged from –3.7 ± 1.6 to 17.1 ± 0.7 °C in February and July, respectively. The mean annual precipitation was 653 ± 37 mm.
| Table 1. The origin of the studied Canadian provenances of lodgepole pine (WGS84). NA – not available. | |||||
| Provenance | Latitude (N) | Longitude (W) | Altitude, m | N of trials | Measurement age |
| Babine Lake | 54°24´ | 125°30´ | 1000 | 2 | 10, 11, 12, 13, 24, 29 |
| Beaton River | 56°50´ | 121°22´ | 800 | 1 | 17 |
| Buick Creek | 56°52´ | 121°30´ | 850 | 2 | 16, 17 |
| Cassior | 58°50´ | 129°25´ | 900 | 2 | 16, 17 |
| Cecil Lake | 56°25´ | 120°00´ | 700 | 3 | 16, 22 |
| Dease Lake | 58°15´ | 130°10´ | 850 | 2 | 16, 17 |
| Ethel Lake | 63°20´ | NA | 900 | 1 | 16 |
| Falkland | 50°32´ | 119°40´ | 1300 | 4 | 12, 16, 22 |
| Fly Hilla | 50°40´ | 119°29´ | 1100 | 2 | 10, 11, 12, 13, 24, 29 |
| Fort Nelson | 58°38´ | 122°41´ | 500 | 2 | 7, 9, 24, 25 |
| Fort St. James | 54°30´ | 124°10´ | 900 | 5 | 12, 16, 18, 22 |
| Francis Lake | 61°10´ | 129°27´ | 950 | 1 | 16 |
| Halfway River | 56°32´ | 122°15´ | 900 | 1 | 17 |
| Hazelton | 55°30´ | NA | 600 | 1 | 16 |
| Hudson Hope | 56°03´ | 121°47´ | 650 | 1 | 22 |
| Jackfish Lake | 63°00´ | NA | 650 | 1 | 16 |
| Lac La Hache | 51°54´ | 121°27´ | 1050 | 2 | 10, 11, 12, 13, 24, 29 |
| Lac le Jeune | 50°29´ | 120°29´ | 1250 | 5 | 12, 16, 18, 22 |
| Lacobie Creek | 52°30´ | 122°00´ | 1000 | 2 | 10, 11, 12, 13, 24, 29 |
| Little Salmon | 62°10´ | NA | 700 | 1 | 16 |
| Mackensia | 58°50´ | 129°25´ | 900 | 1 | 17 |
| Moberly Lake | 55°40´ | 121°25´ | 750 | 5 | 12, 16, 22 |
| Monashee | 50°08´ | 118°30´ | 950 | 4 | 12, 16, 22 |
| Muskey Lake | 55°30´ | NA | 900 | 3 | 16, 22 |
| Nation River | 55°15´ | 124°10´ | 550 | 3 | 10, 11, 12, 13, 18, 24, 29 |
| Nordegg | 52°20´ | 116°25´ | 1400 | 4 | 12, 16, 22 |
| Pink Mountain | 57°00´ | 122°30´ | 850 | 8 | 7, 9, 12, 16, 17, 22, 24, 25 |
| Rocky Mountain House | 52°37´ | 115°20´ | 1000 | 3 | 12, 22 |
| Smithers | 54°15´ | 127°00´ | 650 | 5 | 12, 16, 18, 22 |
| Summit Lake | 54°24´ | 122°37´ | 800 | 2 | 7, 9, 24, 25 |
| Teslin, Yukon | 60°15´ | 132°50´ | 750 | 4 | 12, 16, 22 |
| Tungsten | 61°00´ | 128°50´ | 1050 | 1 | 16 |
| Upper Liard Rivers | 60°10´ | 128°50´ | 700 | 2 | 10, 11, 12, 13, 24, 29 |
| Valemount | 53°00´ | 119°00´ | 850 | 5 | 16, 22, 23 |
| Watson Lake | 60°05´ | 128°37´ | 700 | 2 | 10, 11, 12, 13, 24, 29 |
| Wonowon | 56°36´ | 121°36´ | 850 | 3 | 10, 11, 12, 13, 17, 24, 29 |
The trials were measured at the age of 10–29 years; many of them multiple times. Height (H), stem diameter at breast height (DBH), and diameter of the thickest branch up to 2 m height were measured on each on each tree. The presence of spike knots and animal damage were recorded as binary variables. Stem straightness was evaluated according to three grades (straight, slightly curved, and curved).
2.2 Data analysis
The measurements of DBH and H were scaled (z-scores) by trial to account for the heterogeneity of variance across the trials due to different measurement age (Table 1). Trees with broken tops and leaning stems were excluded from the analysis (2.8% of observations). Random effects models were used to estimate variance components of provenance and provenance × trial interaction of the studied traits. The restricted maximum likelihood approach was used. Analyses were done on tree-level observations. The statistical model in general form was:
![]()
where yijlm – observation (tree), μ – grand mean, pi –effect of provenance i, ptij – effect of interaction between provenance i and trial j, tj – effect of trial j, and b(t)lj– effect of replication l nested within trial j. The influence of individual provenance on the ptij interaction was checked using the leave-one-out approach.
The relationships between latitude, longitude, and altitude of origin (of provenance) and the traits showing a significant provenance variance component were assessed using quadratic two-factor linear regression analysis. The statistical model was:
![]()
where Yij – provenance mean at each trial and age, β0..6 – parameter estimates, X1i, X2j – predictors. The best models (pairs of the best predictors) were selected by the fit statistics (e.g. R2, root mean square error, etc.). Data were analysed in R v. 3.5.1 (R Core Team 2018), using the package lme4 (Bates et al. 2015).
3 Results
The overall 29-year survival of the lodgepole pine was ca. 40%. The overall mean (±confidence interval) H and DBH ranged from 1.36 ± 0.01 to 11.43 ± 0.14 m and from 1.36 ± 0.02 to 12.31 ± 0.25 cm at the age 7 and 29 years, respectively. The provenance and provenance-by-trial variance components of H and DBH were significant (p-value < 0.05) and had a similar magnitude (Table 2), indicating local specifics in the responses of provenances to environment. The probability of wildlife damage showed significant variance by provenance, while the probability of spike knots and survival showed significant interaction variance; yet, the magnitude of the variances were lower than for H and DBH. The provenances had generally similar influence on the interaction, although contribution of Summit Lake was slightly (ca. 5%) higher compared to others, in the case of DBH and H. The among-trial variation was strong for all binomial traits. Stem straightness and branch diameter showed only replication-related variance.
| Table 2. Variance of random effects of provenance, provenance-by-trial, trial, and replication for the studied traits. | |||||||||||||
| Provenance | Provenance × trial | Trial | Replication | Residual variance | |||||||||
| Variance | St. error | p-values | Variance | St. error | p-values | Variance | St. error | p-values | Variance | St. error | p-values | ||
| Continuous (z-scores) | |||||||||||||
| Tree height | 6.85 | 2.62 | <0.01 | 6.14 | 2.48 | <0.01 | - | - | - | 8.98 | 2.99 | <0.01 | 52.66 |
| Stem diameter | 3.67 | 1.92 | <0.01 | 2.42 | 1.56 | <0.01 | - | - | - | 4.05 | 2.01 | <0.01 | 62.17 |
| Diameter of the thickest branch | <0.01 | <0.01 | 0.99 | 0.31 | 0.55 | 0.26 | - | - | - | 1.21 | 1.10 | <0.01 | 53.52 |
| Binomial | |||||||||||||
| Probability of spike knots | 0.01 | 0.10 | 0.54 | 0.04 | 0.18 | <0.01 | 4.28 | 2.07 | <0.01 | 0.06 | 0.24 | <0.01 | - |
| Probability of wildlife damages | 0.12 | 0.35 | 0.01 | 0.02 | 0.13 | 0.54 | 7.20 | 2.68 | <0.01 | 0.55 | 0.74 | <0.01 | - |
| Survival (29 year) | <0.01 | 0.06 | 0.88 | 0.10 | 0.24 | 0.04 | 1.85 | 1.36 | <0.01 | <0.01 | 0.03 | 0.94 | - |
| Multinomial | |||||||||||||
| Stem straightness | <0.01 | 0.02 | 0.54 | <0.01 | <0.01 | 0.99 | <0.01 | 0.08 | 0.02 | 0.01 | 0.07 | <0.01 | - |
The latitude and altitude of the origin of provenance were the best predictors of H (R2 = 0.53), while DBH was best described by latitude and longitude (R2 = 0.46; Fig. 1). The probability of spike knots and wildlife damage were less related to the coordinates of origin (R2 < 0.15). The response surface estimated the best H for provenances from the lower latitudes and intermediate altitudes (50°53´N and 800–1200 m a.s.l., e.g. Fly Hilla, Lac La Hache, and Lacobie Creek), with a distinct decrease under an increasing latitude and altitude (e.g. Dease Lake and Cassior).
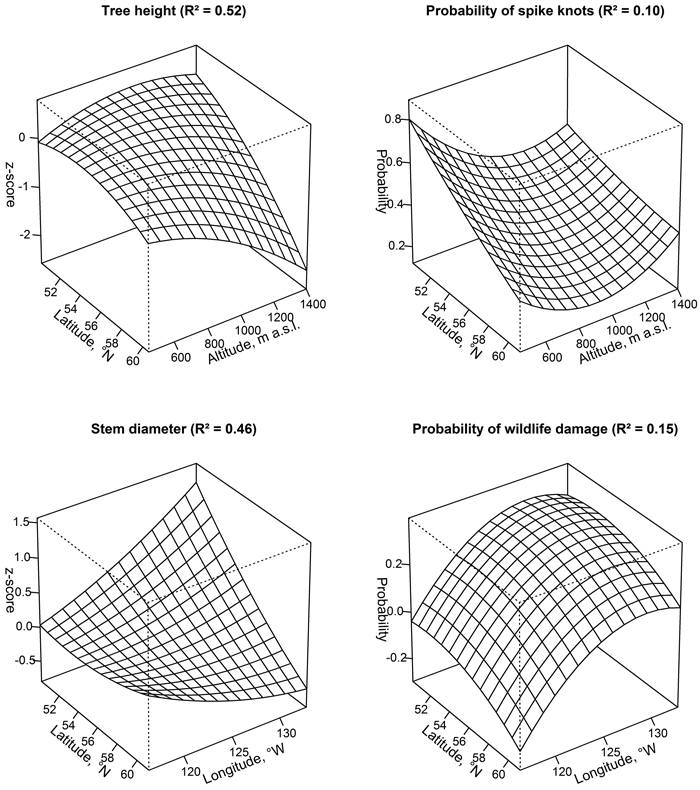
Fig. 1. Predicted tree height, stem diameter at breast height, probability of spike knots, and ungulate damage of the studied lodgepole pine provenances in Latvia according to their origin (longitude, latitude, and altitude). Note that the Y axis differs among the panels.
The largest DBH was associated (Fig. 1) with provenances from low latitudes and high longitudes (50°54´N and >125°W, e.g. Smithers and Babine Lake). Still, the best growing (DBH) provenances were Lacobie Creek, Halfway River, and Fly Hilla (their mean z-score ranging 0.31–0.45, respectively). The probability of spike knots showed the tendency to increase with decreasing latitude, particularly for the lower and higher altitudes. The probability of wildlife damage showed stationary maxima for provenances from mid-latitudes and longitudes, such as Beaton River and Dease Lake, while the Nordegg and Fort Nelson provenances were the less damaged. The most productive Fly Hilla provenance had an intermediate probability of wildlife damage and spike knots (0.15 and 0.37, respectively).
4 Discussion
The overall survival of lodgepole pine, considering self-thinning, was high, suggesting that it might be grown in Latvia. Nevertheless, the significant provenance-by-trial interaction indicated local differences, which were likely due to differing ungulate (e.g. Cervus elaphus L.) population density, hence the intensity of browsing damage (Table 2), which is becoming a more important issue in Eastern Europe (Borkowski et al. 2017). The provenance-by-trial interactions of tree dimensions (Table 2) also suggested effect of site factors, which are significant for trees growing in optimal conditions (Elfving et al. 2001; Düthorn et al. 2013), e.g. under milder climate. Nevertheless, the variation of tree dimensions were affected by provenance (Table 2), indicating the possibility to improve yield by selection among the provenances (Wu and Ying 2004; Bolte et al. 2009). The provenance, hence population related variation of tree dimensions (Table 2) also suggested the potential for breeding (Lindgren 1993; Elfving et al. 2001).
The dimensions of lodgepole pine were related to the geographic location of origin of provenances (Fig. 1), as observed in other studies (Rehfeldt 1987; Lindgren 1993; Elfving et al. 2001; Wu and Ying 2004). Tree height, which is a proxy for productivity (Savill 1997), was superior for the southern provenances from mid-altitudes (Fig. 1), as observed in Germany and Denmark (Larsen and Nielsen 1982; Stephan 1982). Such a response might be explained by an optimum climatic transfer distance comparable to ca. 1–2° degrees of latitude (Rehfeldt 1987), allowing trees to benefit from an altered temperature regime (Jansons et al. 2013a).
The provenance variation in DBH was mainly related to longitude (Fig. 1). This could be explained by the continentality of the origin of provenance. Accordingly, the best performance is expected for trees from the southwestern parts of the distribution, where the climate was milder. This exceeded the considered optimum transfer distance (Rehfeldt 1987), probably due to differences in elevation, indicating the plasticity of secondary growth. However, the predictions of the model (Fig. 1) did not meet any actual provenance (Table 1), suggesting that the effect of latitude and longitude were independent. This might be due to specifics of climate in Northern America as determined by relief and air circulation (Little 1979). The relationship between coordinates of origin and the probability of spike knots and wildlife damage were weaker (Fig. 1), likely due to specialization to local rather than regional conditions (Wu and Ying 2004). Still, the probability of spike knots and wildlife damage was higher for the provenances from lower latitudes, suggesting correlation with productivity (Fig. 1). Alternatively, spike knots might indicate maladaptation to climate (Wu and Ying 2004). Considering that the studied provenances only partially covered the native range of lodgepole pine (Little 1997), the predictions might be slightly biased, and the effect of altitude less pronounced compared to the studies of Rehfeldt (1987) and Ying (1991). Considering optimum transfer distances (Rehfeldt 1987), the results might be extrapolated to the neighbouring Estonia and Lithuania.
Acknowledgements
The study was funded by European Social Fund Project No. 2009/0200/1DP/1.1.1.2.0/09/APIA/VIAA/146 (data collection) European Regional Development Fund Project No 2DP/2.1.1.1/13/APIA/VIAA/031. We acknowledge Julius Danusevičius for constructive ides in data processing.
References
Bates D., Maechler M., Bolker B., Walker S. (2015). Fitting linear mixed-effects models using lme4. Journal of Statistical Software 67(1): 1–48. https://doi.org/10.18637/jss.v067.i01.
Bolte A., Ammer C., Löf M., Madsen P., Nabuurs G.-J., Schall P., Spathelf P., Rock J. (2009). Adaptive forest management in central Europe: climate change impacts, strategies and integrative concept. Scandinavian Journal of Forest Research 24(6): 473–482. https://doi.org/10.1080/02827580903418224.
Borkowski J., Dobrowolska D., Dąbrowski W., Banul R., Załuski D. (2017). Young conifer stands form a deer browsing refuge for an oak admixture: silvicultural implications for forest regeneration under herbivore pressure. European Journal of Forest Research 136(5–6): 787–800. https://doi.org/10.1007/s10342-017-1070-3.
Despain D.G. (2001). Dispersal ecology of lodgepole pine (Pinus contorta Dougl.) in its native environment as related to Swedish forestry. Forest Ecology and Management 141(1–2): 59–68. https://doi.org/10.1016/S0378-1127(00)00489-8.
Düthorn E., Holzkämper S., Timonen M., Esper J. (2013). Influence of micro-site conditions on tree-ring climate signals and trends in central and northern Sweden. Trees 27(5): 1395–1404. https://doi.org/10.1007/s00468-013-0887-8.
Elfving B., Ericsson T., Rosvall O. (2001). The introduction of lodgepole pine for wood production in Sweden. Forest Ecology and Management 141 (1–2): 15–29. https://doi.org/10.1016/S0378-1127(00)00485-0.
Fazekas A.J., Yeh F.C. (2001). Random amplified polymorphic DNA diversity of marginal and central populations in Pinus contorta ssp. latifolia. Genome 44(1): 13–22. https://doi.org/10.1139/g00-082.
IPCC (2013). Climate change 2013: the physical science basis. Cambridge University Press, Cambridge. 1535 p.
Jansons A., Matisons R., Libiete-Zālīte Z., Baders E., Rieksts-Riekstiņš J. (2013a). Relationships of height growth of lodgepole pine (Pinus contorta var. latifolia) and Scots pine (Pinus sylvestris) with climatic factors in Zvirgzde, Latvia. Baltic Forestry 19: 236–244.
Jansons A., Sisenis L., Neimane U., Rieksts-Riekstins J. (2013). Biomass production of young lodgepole pine (Pinus contorta var. latifolia) stands in Latvia. iForest 6: 10–14. https://doi.org/10.3832/ifor0637-006.
Kuznetsova T., Tilk M., Ots K., Lukjanova A., Pärn H. (2009). The growth of lodgepole pine (Pinus contorta var. latifolia Engelm.) in a reclaimed oil shale mining area, abandoned agricultural land and forestland. Baltic Forestry 15: 186–194.
Larsen J.B., Nielsen C.N. (1982). Proveniensforsog med contortafyr (Pinus contorta Dougl.) i Danmark. [Provenance trials with Pinus contorta in Denmark]. Forstlige Forsoegsvaesen i Danmark 38(3): 239–272. [In Danish].
Lindgren D. (1993). Breeding Pinus contorta in different countries. In: Lindgren D. (ed.). Proceedings of Pinus contorta – from untamed forest to domesticated crop. Frans Kempe Symposium, Umeå, August 24–28, 1992.
Lindner M., Maroschek M., Netherer S., Kremer A., Barbati A., Garcia-Gonzalo J., Seidl R., Delzon S., Corona P., Kolström M., Lexer M.J., Marchettie M. (2010). Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. Forest Ecology and Management 259(4): 698–709. https://doi.org/10.1016/j.foreco.2009.09.023.
Little E.L. (1979). Checklist of United States trees (native and naturalized). USDA Agriculture Handbook No. 541, Washington, D.C. 375 p.
Nilsson U., Fahlvik N., Johansson U., Lundström A., Rosvall O. (2011). Simulation of the effect of intensive forest management on forest production in Sweden. Forests 2(1): 373–393. https://doi.org/10.3390/f2010373.
R Core Team (2018). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/.
Rehfeldt G.E. (1987). Components of adaptive variation in Pinus contorta from the Inland Northwest. USDA Forest Service Intermountain Research Station Research Paper INT-375. Moscow, Idaho. 11 p. https://doi.org/10.5962/bhl.title.68869.
Sable I., Grinfelds U., Jansons A., Vikele L., Irbe I., Verovkins A., Treimanis A. (2012). Comparison of the properties of wood and pulp fibers from lodgepole pine (Pinus contorta) and Scots pine (Pinus sylvestris). BioResources 7(2): 1771–1783. https://doi.org/10.15376/biores.7.2.1771-1783.
Savill P., Evans J., Auclair D., Falck J. (1997). Plantation silviculture in Europe. Oxford University Press, Oxford. 308 p.
Schlyter P., Stjernquist I., Bärring L., Jönsson A.M., Nilsson C. (2006). Assessment of the impacts of climate change and weather extremes on boreal forests in northern Europe, focusing on Norway spruce. Climate Research 31(1): 75–84. https://doi.org/10.3354/cr031075.
Stephan B.R. (1982). Herkunftsversuche mit Pinus contorta in der Bundesrepublik Deutschland. [Provenance trials of Pinus contorta in the Federal Republic of Germany]. Forstwissenschaftliches Centralblatt 101(1): 245–259. [In German]. https://doi.org/10.1007/BF02741182.
Wu H.X., Ying C.C. (2004). Geographic pattern of local optimality in natural populations of lodgepole pine. Forest Ecology and Management 194(1–3): 177–198. https://doi.org/10.1016/j.foreco.2004.02.017.
Ying C.C. (1991). Performance of lodgepole pine provenances at sites in southwestern British Columbia. Silvae Genetica 40: 215–223.
Total of 24 references.
