Cheng Bai,
Shixue You,
Weipeng Ku,
Qilin Dai,
Zhengyi Wang,
Mingshui Zhao,
Shuquan Yu 
Life form dynamics of the tree layer in evergreen and deciduous broad-leaved mixed forest during 1996–2017 in Tianmu Mountains, eastern China
Bai C., You S., Ku W., Dai Q., Wang Z., Zhao M., Yu S. (2020). Life form dynamics of the tree layer in evergreen and deciduous broad-leaved mixed forest during 1996–2017 in Tianmu Mountains, eastern China. Silva Fennica vol. 54 no. 2 article id 10167. https://doi.org/10.14214/sf.10167
Highlights
- Biodiversity monitoring was performed over twenty one years in a 1 ha plot
- Life form composition changed significantly between 1996 and 2017
- Evergreen trees in the understory expanded upwards
- Forests were currently dominated by evergreen trees.
Abstract
In the forest areas of eastern China, there is a change from forest dominated by deciduous broad-leaved trees to forest dominated by evergreen broad-leaved trees as the latitude or altitude decreases. Different life forms have different survival strategies to deal with climate change, and studying the life form dynamics of the tree layers in the mixed forest in eastern China, with increasing temperature, can help us understand how the forest responds. This study was performed in a 1 ha plot in evergreen and deciduous broad-leaved mixed forest in Tianmu Mountain National Nature Reserve. Based on the data from two surveys (1996 and 2017), the changes in life form composition and biodiversity over the past 21 years were analyzed. We obtained the following results: (1) The proportion of evergreen trees increased from 55.0% in 1996 to 67.5% in 2017, and the dominance of evergreen species was enhanced. (2) The diversity of both life forms increased, and the tree species were more abundant. (3) The average annual recruitment rate of the evergreen species was 2.1% greater than their mortality rate, and the average annual recruitment rate of the deciduous species was 0.5% less than their mortality rate. (4) The competition among the trees in the small-diameter class (10 cm ≤ DBH < 20 cm) was fierce for many tree species. The proportion of the evergreen species in the small-diameter class was high. The life forms making up the mixed climax forest community has changed over the past 21 years, with the proportion and dominance of evergreen trees increasing significantly.
Keywords
climate change;
biodiversity;
long-term monitoring;
vegetation dynamics
- Bai, State Key Laboratory of Subtropical Silviculture, Zhejiang A&F University, Hangzhou 311300, China E-mail baicheng111@gmail.com
- You, College of Environmental and Resource Sciences, Zhejiang University, Hangzhou 310011, China E-mail sxyou@zju.edu.cn
- Ku, State Key Laboratory of Subtropical Silviculture, Zhejiang A&F University, Hangzhou 311300, China E-mail 2732684475@qq.com
- Dai, State Key Laboratory of Subtropical Silviculture, Zhejiang A&F University, Hangzhou 311300, China E-mail 757692949@qq.com
- Wang, State Key Laboratory of Subtropical Silviculture, Zhejiang A&F University, Hangzhou 311300, China E-mail 237600341@qq.com
- Zhao, Management Bureau of Tianmu Mountain National Nature Reserve, Hangzhou 311311, China E-mail 973659738@qq.com
-
Yu,
State Key Laboratory of Subtropical Silviculture, Zhejiang A&F University, Hangzhou 311300, China
E-mail
yushq@zafu.edu.cn
Received 29 March 2019 Accepted 8 April 2020 Published 27 April 2020
Views 70490
Available at https://doi.org/10.14214/sf.10167 | Download PDF

1 Introduction
Long-term vegetation monitoring has been carried out in different forest types around the world. As global climate change intensifies, ongoing monitoring could be valuable for documenting and understanding forest responses to climate change (Teixeira et al. 2015; Mi et al. 2016). The evergreen and deciduous broad-leaved mixed forest in China’s subtropical mountains is complex and biologically diverse and is considered to be sensitive to changes in climate (Huang et al. 2015a). In this type of forest, at higher latitudes, deciduous trees are dominant in the upper layers, while, evergreen broad-leaved species are less abundant and occupy the lower layer. As latitude decreases, the number of evergreen broad-leaved species increases and they gradually occupy the upper tree layers. The changes in the relative abundance of deciduous and evergreen life forms are often responses to variation in climate (Weng et al. 2017). Thus, data on the distribution of different life forms obtained from long-term monitoring of these forests can be helpful in understanding dynamic changes in forest communities.
The Tianmu Mountains area in eastern China is at a confluence of mid and northern subtropical regions. Since designation as a logging exclusion area in 1956 and a conservation area in 1986, the forest here is protected from human disturbance. The vegetation is very lush, with many well-preserved forests, and high species diversity. Individuals of Cryptomeria fortunei Hooibr. ex Otto & Dietrich, Pseudolarix amabilis (J. Nelson) Rehder and Cyclocarya paliurus (Batalin) Iljinsk. distributed in this area are nationally protected species or are designated as rare and endangered species. In 1996, the Tianmu Mountain National Nature Reserve joined the World Network of Biosphere Reserves of the United Nations Educational, Scientific and Cultural Organization (UNESCO), which has the objective of protecting rare and endangered wildlife as well as forest ecosystems and their biodiversity. Therefore, long-term monitoring of biodiversity dynamics in this nature reserve area is important. In recent years, climate change and its impacts on forest ecosystems have become a hot research topic. A large number of studies on the effects of temperature and precipitation changes on the structure and function of forest ecosystems have been performed in some reserves, and have focused on plant regeneration, species composition, community structure, litter decomposition, and soil respiration (Peng et al. 2009; Teixeira et al. 2013). However, to date, no one has examined the changes in life form and species composition in response to climate change.
Evergreen and deciduous broad-leaved mixed forest is a climax forest community of this mountains area at an altitude of approximately 1000 m. This forest transition zone is a unique natural forest type in China. Plant life form, such as deciduous or evergreen broad-leaved, is a manifestation of the external morphology of plants after adaptation to the environment (Whittaker 1970). Many studies have investigated the spatial distribution, species composition, community regeneration, interspecific relationships, species invasions, rare species distributions (Huang et al. 2015b), dominant populations and their spatial distribution patterns (Shen et al. 2014), and species diversity distribution patterns (Ge et al. 2013; You et al. 2016; Guan et al. 2017; Hu et al. 2018) of evergreen and deciduous broad-leaved mixed forests in this area. However, compared with either evergreen broad-leaved forests or deciduous broad-leaved forests, there have been few studies involving long-term monitoring of broad-leaved mixed forests in subtropical mountains.
In the Tianmu Mountains, with an increase in altitude and the associated drop in temperature, there is a change from a climax forest community dominated by evergreen broad-leaved trees firstly to a community with a balance of evergreen and deciduous broad-leaved trees and then to a community dominated by deciduous broad-leaved trees. The evergreen and deciduous broad-leaved mixed forest in this area is considered to be the zonal climax community at specific elevations (Da et al. 2009). Against the background of climate change in the region, it has become a focus for research to determine whether the distribution of life forms in the region has changed particularly in the complex evergreen and deciduous broad-leaved mixed forest. Therefore, a 1 ha permanent monitoring plot was established at an altitude of 1065 m in 1996 in order to study whether the composition of the forest in terms of life forms would change or not under gradual climate change. In this study, we compared the 1996 and 2017 survey data for the species in the tree layer in the plot to analyze the changes in the species composition and community structure with reference to different life forms over the 21 years. We asked the following questions: (1) How has community structure and species composition of the mid-subtropical mountains evergreen and deciduous broad-leaved mixed forest changed over the past 21 years? (2) How have the dynamics of the life forms changed over this period?
2 Materials and methods
2.1 Study area
The Tianmu Mountain National Nature Reserve is located in the Lin’an District, Hangzhou City, Zhejiang Province, China (30°18′–30°25´N, 119°24′–119°28´E) and has an area of 4284 ha. The altitude of the area is 300–1506 m and is in a transition zone between the mid-subtropical zone and the northern subtropical zone. The average annual temperature at the top of the mountains (Xianrending weather station is at an altitude of 1506 m) is 9.0 °C and the average annual precipitation is 1751 mm (Da et al. 2009). The vegetation types are rich and diversified, and the vegetation distribution shows obvious vertical changes. Evergreen broad-leaved forest is a typical zonal vegetation that is mainly distributed at an altitude below 850 m, evergreen and deciduous broad-leaved mixed forest is the main vegetation type, which is mainly distributed at the altitude of 850–1100 m, deciduous broad-leaved forest is distributed at the altitude of 1100–1380 m, and deciduous coppice is distributed at the altitude above 1380 m. The floristic composition of the study area is complex with 1718 species of seed plants including 35 key national preserved species accounting for 64% of the key preserved species in the Zhejiang Province, and 171 species of ferns as one of the regions with relatively well-preserved vegetation in eastern China.
2.2 Sample plot and survey methods
To monitor the long-term dynamics of the evergreen and deciduous broad-leaved mixed forest, a 1 ha (100 m × 100 m) plot was identified in the Tianmu Mountain National Nature Reserve in 1996. The center of the plot is located at 30°34´N, 119°43´E at an altitude of 1065 m. In 1996, all woody plants with a diameter at breast height (DBH) of 10 cm or more were numbered and tagged, and names of species, heights, DBHs, crown widths, branch heights, growth state, and position coordinates were recorded. The site of DBH measurement was marked with paint. In the absence of human interference, the plots were remeasured in 2017, following the same survey method as used in 1996. The temperature and precipitation data in this paper were obtained from the monitoring data of the Hangzhou Ground Weather Station in Hangzhou, China which is about 80 km from our study site.
2.3 Methods of data processing and analysis
In this study, the data of the two surveys conducted in 1996 and 2017 were analyzed. The data of the plants with DBH of ≥10 cm (excluding vines) in the tree layer were used to compare and analyze the life form composition and dynamics of the tree species in the community.
The life forms of the tree layer in this study area were divided into two types, evergreen and deciduous (Givnish et al. 2002) depending on the presence of leaves in the tree canopy throughout the entire year.
The equation for the calculation of the importance value (IV) of tree species was:
![]()
The equations for the calculation of mortality (M) and recruitment rate (R) are as follows:

where F0 is the number of trees of a species in the first survey, Ft is the number of trees of this population in the second survey, St is the number of the surviving trees of the population at the second survey, and T is the interval time between the two surveys.
The cumulative mortality rate of the different life forms was calculated as the number of deaths divided by the original number of trees, and the cumulative recruitment rate was calculated as the number of new trees divided by the number of surviving trees. Hill numbers (Chao et al. 2012) were used to measure diversity of tree species, including species richness (N0), exponential of Shannon entropy (N1), and the inverse Simpson concentration (N2). Species with ≤1 tree per ha were defined as rare species, species with 1−10 trees per ha were defined as an occasional species, and species with >10 trees per ha were defined as common species (Hubbell and Foster 1986). The trees in the community were divided into six DBH classes at intervals of 10 cm: 10 ≤ DBH < 20, 20 ≤ DBH < 30, 30 ≤ DBH < 40, 40 ≤ DBH < 50, 50 ≤ DBH < 60, and DBH ≥ 60 cm. And division of the height was divided into layer C (H < 10 m), layer B (10 ≤ H < 20 m), and layer A (H ≥ 20 m) (Lu et al. 2013).
In order to eliminate the random drift of the data between the two surveys, we divided the data into 25 observations (20 m × 20 m). Paired sample t-tests were used to analyze the floristic differences between evergreen and deciduous species between 1996 and 2017. Data processing, analysis, and graphing were performed in R software 3.5.0 and Origin 2017.
3 Results
3.1 Changes in species composition
The analysis of paired sample t-test showed that the number of evergreen trees increased significantly, and the number of deciduous trees decreased significantly. The species composition changed significantly during the past 21 years (Table 1). For example, in 1996, the proportions of evergreen and deciduous trees in the community were 55.0% and 45.0% respectively, while they were 67.5% and 32.5% in 2017, respectively. The basal area of the evergreen trees showed an upward trend, while the basal area of the deciduous trees showed a downward trend. From 1996 to 2017, the density of evergreens increased by 2.19 m2 ha–1 (11.1%), and that of the deciduous trees decreased by 0.66 m2 ha–1 (3.6%). The proportion of the importance value of the evergreen trees increased from 49.2% to 56.8%, in the meantime, deciduous trees decreased from 50.8% to 43.2% (Table 2).
| Table 1. Result of Paired samples t-test for quantity of evergreen and deciduous trees between 1996 and 2017, in Tianmu Mountains, eastern China. | |||||
| Life forms | mean | se | t | df | p |
| Evergreen1996 – Evergreen2017 | –6.56 | 1.41 | –4.65 | 24 | 0.00 |
| Deciduous1996 – Deciduous2017 | 0.96 | 0.35 | 2.75 | 24 | 0.01 |
| Table 2. Changes of the evergreen and deciduous species in Tianmu Mountains, eastern China, in 1996 and 2017. | |||||||||||
| Life forms | Stem density (stems ha–1) | Basal area (m2 ha–1) | IV | Recruit trees | Dead trees | Survival trees | Recruitment rate | Mortality rate | |||
| 1996 | 2017 | 1996 | 2017 | 1996 | 2017 | 1996–2017 | 1996–2017 | 1996–2017 | 1996–2017 | 1996–2017 | |
| Evergreen | 304 | 468 | 19.80 | 21.99 | 49.22 | 56.84 | 268 | 104 | 201 | 4.0% | 1.9% |
| Deciduous | 249 | 225 | 18.36 | 17.70 | 50.78 | 43.16 | 90 | 114 | 134 | 2.5% | 2.9% |
| Total | 553 | 693 | 38.16 | 39.69 | 100 | 100 | 358 | 218 | 335 | ||
3.2 Species recruitment and mortality
Evergreen trees showed a high recruitment rate and a low mortality rate. The cumulative recruitment rate of the deciduous trees was lower than the cumulative mortality rate and the number of trees decreased. 268 new evergreen trees and 90 deciduous trees were recruited over the 21-year period, with an average annual recruitment rate of 4.0% and 2.5%, respectively. 103 evergreen trees and 115 deciduous trees died, with an average annual mortality rate of 1.9%, 2.9%, respectively. (Table 2).
Between 1996 to 2017, recruitment rate differed among the 8 evergreen tree species, and the cumulative recruitment rate was 134.0%. Seven evergreen tree species were observed to have dead or dying trees, and the cumulative mortality rate of the trees was 34.2%. One evergreen species left the tree layer while another one entered. Similarly, recruitment rate differed among the 36 deciduous tree species, and the cumulative recruitment rate was 66.7%. A total of 24 deciduous tree species had dead or dying trees, and the cumulative mortality rate of the trees was 45.8%. Five deciduous species left and 14 species entered the tree layer.
3.3 Changes in species diversity
Over the 21 years the diversity indices of both the evergreen and deciduous trees showed an upward trend, except for N2. For the evergreen species, the N0 remained unchanged, N1 increased from 8.76 to 10.59 (rate change, 22.1%) and N2 declined from 1.47 to 1.32 (rate change, 10.2%). For the deciduous tree species, the N0 increased from 31 to 40 (rate change, 29.0%), N1 increased from 48.32 to 58.52 (rate change, 21.1%) and N2 declined from 1.16 to 1.14 (rate change, 1.7%) (Table 3).
| Table 3. Diversity indices (Hill Numbers: N0, N1, N2) of the evergreen and deciduous life forms in 1996 and 2017. | ||||||
| Life forms | N0 | N1 | N2 | |||
| 1996 | 2017 | 1996 | 2017 | 1996 | 2017 | |
| Evergreen | 9 | 9 | 8.67 | 10.59 | 1.47 | 1.32 |
| Deciduous | 31 | 40 | 43.38 | 58.52 | 1.16 | 1.14 |
3.4 Changes in species height
The evergreen trees in all layers showed an increasing trend, and the deciduous trees had a decreasing trend. Both in 1996 and 2017, N0 of the tree species in layer B and layer C was high, whereas N0 in layer A was relatively low. Furthermore, N0 in each layer of the evergreens was lower than that of the deciduous trees. From 1996 to 2017, among the evergreens, only one species was added in layer C, while among the deciduous trees, 2, 8 and 2 species were added in layer A, layer B and layer C, respectively, most of which were occasional species (Table 4).
| Table 4. Height structure of tree layer in Tianmu Mountains, eastern China, in 1996 and 2017. | |||||||
| Layer | Life forms | Species | Trees | Net gains | Mortality | ||
| 1996 | 2017 | 1996 | 2017 | 1996–2017 | 1996–2017 | ||
| A | Evergreen | 4 | 4 | 22 | 26 | 4 | 1 |
| Deciduous | 9 | 11 | 66 | 65 | –1 | 15 | |
| B | Evergreen | 8 | 8 | 76 | 144 | 68 | 24 |
| Deciduous | 22 | 30 | 144 | 93 | 51 | 59 | |
| C | Evergreen | 7 | 8 | 206 | 298 | 92 | 79 |
| Deciduous | 24 | 26 | 69 | 67 | –2 | 40 | |
Regarding the number of trees, in 1996, the evergreen trees were mainly distributed in layer C, and the deciduous trees were dominant in layers A and B. While in 2017, evergreen life forms were dominant in the layers B and C, and deciduous trees mostly existed in the layer A (Table 4). From 1996 to 2017, the proportion of the evergreens in all the layers showed an upward trend. The cumulative number of evergreen trees in layer A increased by 23.8%, but the death rate was only 4.6%. While, the cumulative number of deciduous trees was increased by 27.4%, and the death rate was 22.7%. In layer B, the proportion of the evergreens exceeded that of the deciduous trees (Table 4). Although the number of dead deciduous trees was much higher than that of the evergreen, the net growth of the evergreens was higher than that of the deciduous trees. Note that Layer C had the highest proportion of trees. The total number of evergreen and deciduous trees increased by 134.7% and 131.0%, and the death rate was 38.4% and 58.0%, respectively.
3.5 Changes in species diameter classes
Based on the number of trees in each diameter class, the number of trees with a small diameter was large, and the proportion of the evergreen trees increased. As the diameter increased, the number of trees decreased sharply. In trees with a DBH of > 50 cm, the number of trees tends to be stable, the diameter structure of the community showed an inverted “J” shape. As shown in Fig. 1, the number of trees with a small diameter (10 cm ≤ DBH < 20 cm) in the community was the highest. In 1996, the evergreen and deciduous trees accounted for 63.5% and 36.5%, respectively, they were 77.9% and 22.1% in 2017. The trees with a large diameter (DBH ≥ 60 cm) always maintained a proportion of 50% of each life form.
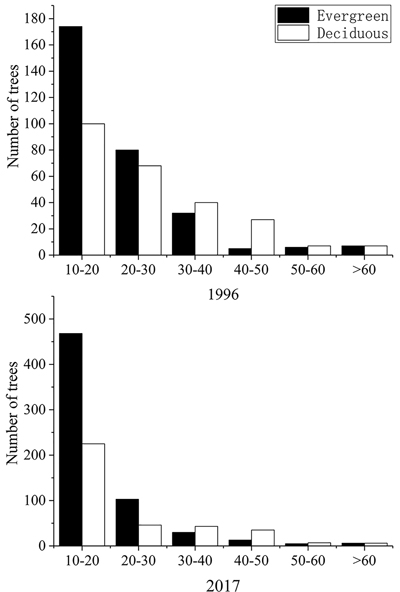
Fig. 1. Size distribution of the evergreen and deciduous trees. in Tianmu Mountains, eastern China, in 1996 and 2017.
In terms of recruited and dead trees in different diameter classes, the number of trees in the small-diameter class was the largest (Fig. 2). In general, the number of new evergreen trees entering the tree layer with a small diameter was 2.84 times that of the deciduous trees. The mortality rate of the deciduous trees is higher than that of the evergreens, and the competition among the trees in the small-diameter class was the most intense with a high mortality rate.
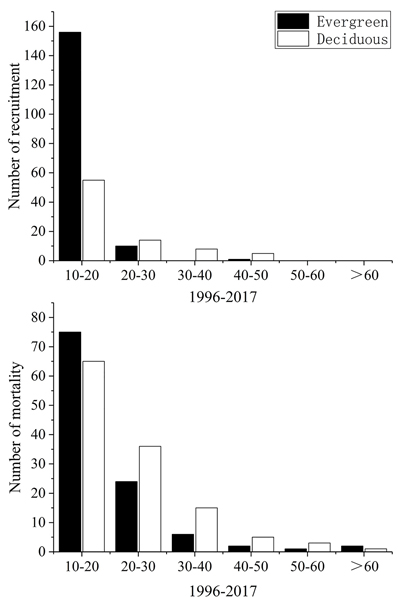
Fig. 2. Size distribution of recruits and dead trees for each life form in Tianmu Mountains, eastern China, in 1996 and 2017.
3.6 Analysis of the major tree species
There were the ten tree species having a total importance value of >70% (Table 5), playing key roles in determining the nature and structural characteristics of the community. In the past 21 years, the importance values of these dominant tree species in the community remained stable. The importance values of Daphniphyllum macropodum Mig. and Lithocarpus brevicaudatus (Skan) Hayata increased significantly, and that of C. paliurus decreased slightly. Among the evergreens, 162 D. macropodum trees showed a high net increase (accounting for 45.3% of the total recruitment of the community), and their importance value increased from 3.2% to 14.7%. The rank of D. macropodum in the tree species rose from 12 to 1, making it the dominant tree species in the community. L. brevicaudatus also maintained a high recruitment rate and a low mortality rate, and the number of trees increased significantly, and the importance value also increased. Among the deciduous trees, the recruitment rate of C. paliurus was relatively low owing to its relatively high mortality rate resulting in a significant decrease in the importance value (Table 5).
| Table 5. Floristic changes of the top ten tree species ranked by importance values, in Tianmu Mountains, eastern China, in 1996 and 2017. | ||||||||
| Species name | IV | Stem density (stems ha–1) | Mortality rate 1996–2017 | Recruitment rate 1996–2017 | Life forms | |||
| 1996 | 2017 | 1996 | 2017 | |||||
| 1 | Daphniphyllum macropodum | 3.18 | 14.65 | 15 | 177 | 5.2% | 16.9% | Evergreen |
| 2 | Cyclobalanopsis myrsinifolia (Blume) Oerst. | 17.37 | 10.86 | 153 | 118 | 2.1% | 0.8% | Deciduous |
| 3 | Cunninghamia lanceolata (Lamb.) Hook. | 8.98 | 10.08 | 64 | 64 | 0.6% | 0.6% | Evergreen |
| 4 | Cyclocarya paliurus | 13.4 | 9.69 | 80 | 56 | 3.3% | 1.6% | Deciduous |
| 5 | Liquidambar acalycina H.T. Chang | 7.66 | 7.85 | 31 | 33 | 0.0% | 0.3% | Deciduous |
| 6 | Lithocarpus brevicaudatus | 3.07 | 6.41 | 22 | 52 | 2.5% | 6.6% | Evergreen |
| 7 | Cryptomeria fortunei | 8.98 | 6.36 | 10 | 9 | 1.1% | 0.6% | Evergreen |
| 8 | Litsea auriculata S.S. Chien & W.C. Cheng | 3.53 | 3.64 | 15 | 15 | 0.7% | 0.7% | Deciduous |
| 9 | Dendrobenthamia japonica var. chinensis (Osborn) W.P.Fang | 2.66 | 3.18 | 16 | 25 | 2.7% | 4.9% | Deciduous |
| 10 | Eurya hebeclados Ling | 3.33 | 3.09 | 26 | 28 | 5.6% | 5.9% | Evergreen |
4 Discussion
4.1 Changes in the floristic characteristics of the different life forms
In response to different types of climate changes, evergreen and deciduous trees usually exhibit different ecological strategies (van Ommen Kloeke et al. 2012). Since the establishment of Tianmu Mountain National Nature Reserve in 1956, protection measures have been adopted to better protect the vegetation, especially the tree layer, from human disturbance. In 1996, the dominant deciduous trees in the canopy significantly affected the forest understory sunlight condition, affecting forest regeneration and species diversity (Quigley et al. 2003). During the period of leaf-off (from late autumn to early spring), the amount of sunlight in the understory increases, but the low winter temperature may limit the growth of evergreen trees in the understory of the forest (Wyka and Oleksyn 2014). Thus, this kind of competition eventually formed a pattern of deciduous broad-leaved trees dominating in the canopy and the evergreen trees dominating in the understory of the forest. According to the meteorological data in this area, the average annual temperature (Fig. 3 A) and the coldest and hottest monthly average temperature (Figs. 3 C, D) have all increased, and the annual precipitation (Fig. 3 B) remain stable in the past 21 years. With an increase in temperature, the growth season of the evergreen broad-leaved tree species was longer than that of the deciduous broad-leaved trees. The growth rate of the evergreen broad-leaved species was higher than that of deciduous broad-leaved species, so the deciduous species were gradually excluded from the competition (Woodward 1990). In our study area, tree layer changed from a relatively balanced state between the evergreen and deciduous trees to a state of evergreen tree dominance. Therefore, we speculate that the changes in the proportion of evergreen and deciduous trees is a reflection of temperature changes.
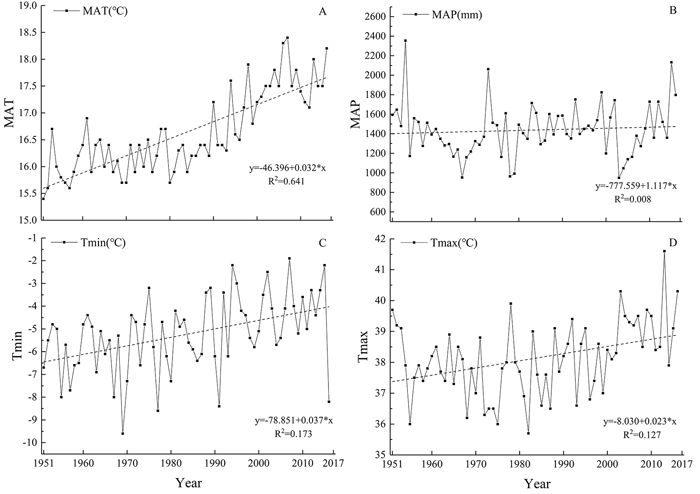
Fig. 3. Changes in annual temperature (MAT), annual precipitation (MAP), annual hottest month average temperature (Tmax) and coldest month average temperature (Tmin) of 60 years in Tianmu Mountains, eastern China. View larger in new window/tab.
4.2 Analysis of recruitment and mortality
Forests are dynamic communities, and the dynamic changes in the mortality and recruitment rate could lead to dynamic changes at the plant individual, population, and community levels, which can significantly alter the structure and life form composition of the forest stand (Acker et al. 2014). Moreover, climate warming and drought have been identified as the major driving factors that may change the life form composition of a forest community (Allen et al. 2010). In our study, most of the tree species showed different degrees of death and recruitment over 21 years. The number of evergreen tree species increased, such as D. macropodum and L. brevicaudatus, while deciduous tree species with a large DBH in the top layer die. Owing to the imbalance between mortality and recruitment rates, the dominance of the shade-tolerant evergreen species in the community was enhanced, whereas the deciduous species were largely eliminated from the community. Intense understory competition also has occurred in evergreen broad-leaved forests (Wang et al. 2001). According to the plant death structure of each diameter class, both evergreen and deciduous tree species had an inverted “J” type distribution. Fig. 2 shows that the mortality of the trees with small DBH was the highest and with medium DBH was the smallest, which is consistent with the findings obtained from the evergreen and deciduous broad-leaved mixed forest in the Karst region of southwest China (Liu et al. 2018) and the Tiantong National Forest Park (Wu et al. 2014). Note that the trees with small DBH also showed the highest recruitment rate. While, in the medium- and large-diameter classes, the recruitment of the trees was mainly due to the growth of the surviving deciduous tree species as they entered the large diameter classes. This forest community is mainly composed of deciduous trees, and hence, the deciduous trees may suffer more losses (Mori et al. 2007). Furthermore, the forest opening formed after canopy destruction is beneficial for the growth of the evergreen trees with small diameter. In our surveys, some local fallen tree areas happened to provide a living space for the small and medium trees belonging to some occasional species.
4.3 Changes in the species diversity
Species diversity reflects the richness of the species, the stability and dynamics of the community, and the relationships among the different natural geographical conditions and the community (Leinster et al. 2012). In these two surveys, the diversity of both the evergreen and deciduous trees showed an increasing trend. N0 of the evergreens was unchanged, whereas that of the deciduous trees increased and most of these new species were occasional species. Some studies have also found a lot of occasional species in the understory of evergreen broad-leaved forest community (Ding et al. 2015) and montane rainforest areas (Zhang et al. 2015). The richness of the understory tree species had changed most significantly (Huelbert et al. 2006). Occasional species play an important role in maintaining diversity of tree layer in forest communities, and that is also the reason why the diversity of deciduous tree species increased but the stem density decreased. In terms of the stem density, the evergreen trees increased and the deciduous species gradually decreased, resulting that the evergreen species were continuously invading and became dominant in the community. In the end, the community tends to become an evergreen broad-leaved forest which is the zonal vegetation in this area. Vegetation dynamics is strongly influenced by the history of land use, climate, and soil (Madeira et al. 2009). Although the area of this study is a nature reserve without human interference, the diversity of evergreen and deciduous species here was changed. Thus, it can be a response of two different life forms to habitat changes in the study area.
5 Conclusion
In general, in the backdrop of continuous warming, the life form composition of the tree layer of the evergreen and deciduous broad-leaved mixed forest in the Tianmu Mountains was changed. The dominance of the evergreen component increased, and the appearance of the tree layer of the community developed from a relatively balanced state of evergreen and deciduous species towards an evergreen-dominated state. In the background of further extreme warming (Sévellec et al. 2018), the vegetation dynamics of the study area need to be further explored. Observation for a longer time scale and monitoring of different altitudes and latitude gradients can enrich the data sources and help us further understand the evolution of the forest ecosystems. Future research can also increase the correlation analysis of multi-climatic environmental factors and vegetation change response and provide theoretical support for the sustainable development and management of forest ecosystems based on richer and more rigorous statistical analysis results.
Acknowledgments
This study was funded by Long-term monitoring plot construction project of Tianmu Mountain National Nature Reserve; Suitable environment and high-efficiency compound management technology research project of main rare tree species in Zhejiang Province, China (Grant Nos.2017C02028). We thank Dr. Quan Li and Mr. Chao Zhang for their help in the statistical analyses. We also thank the journal editors and two anonymous reviewers for their constructive comments on the manuscript.
References
Acker S., Boetsch J., Bivin M., Whiteaker L., Cole C., Philippi T. (2015). Recent tree mortality and recruitment in mature and old-growth forests in western Washington. Forest Ecology and Management 336: 109–118. https://doi.org/10.1016/j.foreco.2014.10.008.
Allen C., Macalady A., Chenchouni H., Bachelet D., Mcdowell N., Vennetier M., Kitzberger T., Rigling A., Breshears D., Hogg E., Gonzalez P., Fensham R., Zhang Z., Castro J., Demidova N., Lim J., Allard G., Running S., Semerci A., Cobb N. (2010). A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest Ecology and Management 259(4): 660–684. https://doi.org/10.1016/j.foreco.2009.09.001.
Anderson-Teixeira K.J., Miller A.D., Mohan J.E., Hudiburg T.W., Duval B.D., DeLucia E.H. (2013). Altered dynamics of forest recovery under a changing climate. Global Change Biology 19(7): 2001–2021. https://doi.org/10.1111/gcb.12194.
Anderson-Teixeira K.J., Davies S.J., Bennett A.C., Gonzalez-Akre E.B., Muller-Landau H.C., Wright S.J., Salim K.A., Zambrano A.M., Alonso A., Baltzer J.L., Basset Y., Bourg N.A., Broadbent E.N., Brockelman W.Y., Vejchewin S.B., Burslem D.F.R.P., Butt N., Cao M., Cardenas D., Chuyong G.B., Clay K., Cordell S., Dattaraja H.S., Deng X., Detto M., Du X., Duque A., Erikson D..L, Ewanco C.E.N., Fischer G.A., Fletcher C., Foster R.B., Giardina C.N.P., Gilbert G.S., Gunatilleke N., Gunatilleke S., Hao Z., Hargrove W.W., Hart T.A., Hau B.C.H., He F., Hoffman F.M., Howe R.W., Hubbell S.P., Inman-Narahari F.M., Jansen P.A., Jiang M., Johnson D.J., Kanzaki M., Kassim A.R., Kenfack D., Kibet S., Kinnaird M.F., Korte L., Kral K., Kumar J., Larson A.J., Li Y., Li X., Liu S., Lum S.K.Y., Lutz J.A., Ma K., Maddalena D.M., Makana J.-R., Malhi Y., Marthews T., Serudin R.M., McMahon S.M., McShea W.J., Memiaghe H.R., Mi X., Mizuno T., Morecroft M., Myers J.M., Novotny V., Oliveira A.A. de, Ong P.S., Orwig D.A., Ostertag R., Ouden J.D., Parker G.G., Phillips R.P., Sack L., Sainge M.N., Sang W., SRI-Ngernyuang K., Sukumar R., Sun I.-F., Sungpalee W., Suresh H.S., Tan S., Thomas S.G., Thomas D.W., Thompson J., Turner B.L., Uriarte M., Valencia R., Vallejo M.I., Vicentini A., Vrška T., Wang X., Wang X., Weiblen G., Wolf A., XU H., Yap S., Zimmerman J. (2015). CTFS-ForestGEO: a worldwide network monitoring forests in an era of global change. Global Change Biology 21(2): 528–549. https://doi.org/10.1111/gcb.12712.
Chao A., Chiu C.-H., Hsieh T.C. (2012). Proposing a resolution to debates on diversity partitioning. Ecology 93(9): 2037–2051. https://doi.org/10.1890/11-1817.1.
Condit R., Sukumar R., Hubbell S.P., Foster R.B. (1998). Predicting population trends from size distributions: a direct test in a tropical tree community. American Naturalist 152(4): 495–509. https://doi.org/10.1086/286186.
Da L., Kang M., Song K., Shang K., Yang Y., Xia A., Qi Y. (2009). Altitudinal zonation of human-disturbed vegetation on Mt. Tianmu, eastern China. Ecological Research 24: 1287–1299. https://doi.org/10.1007/s11284-009-0613-6.
Ding H., Yang Y., Xu H., Fang Y., Chen X., Yang Q., Yi X., Xu H., Wen X., Xu X. (2015). Species composition and community structure of the typical evergreen broad- leaved forest in the Wuyi Mountains of southeastern China. Acta Ecologica Sinica 4: 1142–1154. [In Chinese]. https://doi.org/10.5846/stxb201305050924.
Ge J., Xiong G., Zhao C., Shen G., Xie Z. (2013). Short-term dynamic shifts in woody plants in a montane mixed evergreen and deciduous broadleaved forest in central China. Forest Ecology and Management 310: 740–746. https://doi.org/10.1016/j.foreco.2013.09.019.
Givnish T.J. (2002). Adaptive significance of evergreen vs. deciduous leaves: solving the triple paradox. Silva Fennica 36(3): 703–743. https://doi.org/10.14214/sf.535.
Guan J., Shang T., Yi L., Ye N., Yu S. (2017). Biomass change and community succession characteristics of dominant species in evergreen and deciduous broad-leaved mixed forests in Tianmu Mountains. Acta Ecologica Sinica 37: 6761–6772. [In Chinese]. https://doi.org/10.5846/stxb201605070878.
He F., Legendre P., Lafrankie J.V. (2012). Distribution patterns of tree species in a Malaysian tropical rain forest. Journal of Vegetation Science 8(1): 105–114. https://doi.org/10.2307/3237248.
Hu F., Zeng F., Du H., Peng W., Zhang F., Tan W., Song T. (2018). Scale-dependent spatial patterns for species diversity in a karst evergreen and deciduous broad-leaved mixed forest of northwest Guangxi. Acta Ecologica Sinica 38: 6074–6083. [In Chinese]. https://doi.org/10.5846/stxb201709231713.
Huang Y., Ai X., Yao L., Zang R., Dang Y., Huang J., Feng G., Liu J. (2015a). Changes in the diversity of evergreen and deciduous species during natural recovery following clear-cutting in a subtropical evergreen-deciduous broadleaved mixed forest of central China. Tropical Conservation Science 8(4): 1033–1052. https://doi.org/10.1177/194008291500800413.
Huang Y., Yao L., Ai X., Lü S., Ding Y. (2015b). Quantitative classification of the subtropical evergreen-deciduous broadleaved mixed forest and the deciduous and evergreen species composition structure across two national nature reserves in the southwest of Hubei, China. Chinese Journal of Plant Ecology 39(10): 990–1002. [In Chinese]. https://doi.org/10.17521/cjpe.2015.0096.
Hubbell S., Foster R. (1986). Commonness and rarity in a neotropical forest: implications for tropical tree conservation. Plant Ecology 8: 205–231.
Huelber K., Gottfried M., Pauli H., Reiter K., Winkler M., Grabherr G. (2006). Phenological responses of snowbed species to snow removal dates in the central alps: implications for climate warming. Arct. Antarctic and Alpine Research 38(1): 99–103. https://doi.org/10.1657/1523-0430(2006)038[0099:PROSST]2.0.CO;2.
Leinster T., Cobbold C.A. (2012). Measuring diversity: the importance of species similarity. Ecology 93(3): 477–489. https://doi.org/10.1890/10-2402.1.
Liu L., Jian N., Zhong Q., Gang H., Zhang Z. (2018). High mortality and low net change in live woody biomass of Karst evergreen and deciduous broad-leaved mixed forest in southwestern China. Forests 9(5) article 263. https://doi.org/10.3390/f9050263.
Lou T., Zhao M., Yang S., Pang C., Wang Z., Liu L. (2004). Ancient and famous tree resources in Tianmu Mountain national nature reserve. Journal of Zhejiang Forestry Collage 23: 269–274. [In Chinese].
Lu Z., Bao D., Guo Y., Lu J., Wang Q., He D., Zhang K., Xu Y., Liu H., Meng H., Huang H., Wei X., Liao J., Qiao X., Jiang M., Gu Z., Liao C. (2013). Community composition and structure of badagongshan (BDGS) forest dynamic plot in a mid-subtropical mountain evergreen and deciduous broad-leaved mixed forest, central China. Plant Science Journal 31(4): 336–344. https://doi.org/10.3724/SP.J.1142.2013.40336.
Madeira B.G., Nunes Y., Espírito-Santo M.M., Neto S.D., Nunes Y.R.F., Sánchez Azofeifa G.A., Fernandes G.W., Quesada M. (2009). Changes in tree and liana communities along a successional gradient in a tropical dry forest in south-eastern Brazil. Plant Ecology 201: 291–304. https://doi.org/10.1007/s11258-009-9580-9.
Magurran A.E. (1988). Ecological diversity and its measurement. Springer, Dordrecht. https://doi.org/10.1007/978-94-015-7358-0.
Mi X., Guo J., Hao Z., Xie Z., Guo K., Ma K. (2016). Chinese forest biodiversity monitoring: scientific foundations and strategic planning. Biodiversity Science 24(11): 1203–1219. https://doi.org/10.17520/biods.2015313.
Mori A.S., Mizumachi E., Komiyama A. (2007). Roles of disturbance and demographic non-equilibrium in species coexistence, inferred from 25-year dynamics of a late-successional old-growth subalpine forest. Forest Ecology and Management 241(1–3): 74–83. https://doi.org/10.1016/j.foreco.2006.12.031.
Paltt W.J., Quigley M.F. (2003). Composition and structure of seasonally deciduous forests in the Americas. Ecological Monographs 73(1): 87–106. https://doi.org/10.1890/0012-9615(2003)073[0087:CASOSD]2.0.CO;2.
Peng S., Piao S., Wang T., Sun J., Shen Z. (2009). Temperature sensitivity of soil respiration in different ecosystems in China. Soil Biology and Biochemistry 41(5): 1008–1014. https://doi.org/10.1016/j.soilbio.2008.10.023.
Sévellec F., Drijfhout S.S. (2018). A novel probabilistic forecast system predicting anomalously warm 2018–2022 reinforcing the long-term global warming trend. Nature Communications 9 article 3024. 12 p. https://doi.org/10.1038/s41467-018-05442-8.
Shen M., Zhang C., Zheng C., Zhang X., Yi L., Yu S. (2014). Dominant species populations and their spatial patterns in evergreen and deciduous broad-leaved forest in Tianmu Mountains, China. Journal of Zhejiang University 41: 715–724. [In Chinese].
van Ommen Kloeke A.E.E., Douma J.C., Ordoñez J.C., Reich P.B., van Bodegom P.M. (2012). Global quantification of contrasting leaf life span strategies for deciduous and evergreen species in response to environmental conditions. Global Ecology and Biogeography 21(2): 224–235. https://doi.org/10.1111/j.1466-8238.2011.00667.x.
Wang Y., Mi X., Chen S., Li M., Yu M. (2011). Regeneration dynamics of major tree species during 2002–2007 in a sub-tropical evergreen broad-leaved forest in Gutianshan national nature reserve in East China. Biodiversity Science 19(2): 178–189. https://doi.org/10.3724/SP.J.1003.2011.12244.
Wasingya E., Amani C., Lejoly J., Nshimba H. (2018). Diversity, structure and effect of rare species and relative abundance of the over-represented species on arborescent layers’ diversity. Journal of Biology and Environment Science 13: 102–112.
Weng E., Farrior C.E., Dybzinski R., Pacala S.W. (2017). Predicting vegetation type through physiological and environment interactions with leaf traits: evergreen and deciduous forests in an earth system modeling framework. Global Change Biology 23(6): 2482–2498. https://doi.org/10.1111/gcb.13542.
Whittaker R.H. (1970). Communities and ecosystems. Macmillan, New York. 162 p.
Woodward F.I. (1990). The impact of low temperatures in controlling the geographical distribution of plants [and Discussion]. Philosophical Transactions of the Royal Society of London series B-biological Sciences 326(1237): 585–593. https://doi.org/10.1098/rstb.1990.0033.
Wu Y., Guo C., Ni J. (2014). Dynamics of major forest vegetations in Tiantong national forest park during the last 30 years. Chinese Journal of Applied Ecology 25: 1547–1554.
Wyka T., Oleksyn J. (2014). Photosynthetic ecophysiology of evergreen leaves in the woody angiosperms – a review. Dendrobiology 72: 3–27. https://doi.org/10.12657/denbio.072.001.
You S., Zhang C., Ku W., Zhu H., Zhao M., Yu S. (2016). Community dynamics of arborous layer in the evergreen and deciduous broad-leave mixed forests during 1996–2012 in Tianmu Mountains. Scientia Silvae Sinicae 52: 1–9. [In Chinese].
Yue H., Jin Z., Shao W., Jiang J., Xia X., Huang L. (2012). Structures and dynamics of Castanopsis sclerophylla population in Tianmu Mountains of Zhejiang Province. Bulletin of Botanical Research 32: 473–480.
Zhang G., Du F., Wang H., Li M., Meng F. (2015). Study on tree layer dynamic in Xishuangbanna rain forest based on 20 years monitoring. Acta Ecologica Sinica 35: 4053–4062. [In Chinese]. https://doi.org/10.5846/stxb201311082694.
Total of 40 references.
Send to email
