Ulf Sikström  ,
Hannu Hökkä
,
Hannu Hökkä
Interactions between soil water conditions and forest stands in boreal forests with implications for ditch network maintenance
Sikström U., Hökkä H. (2015). Interactions between soil water conditions and forest stands in boreal forests with implications for ditch network maintenance. Silva Fennica vol. 50 no. 1 article id 1416. https://doi.org/10.14214/sf.1416
Highlights
- Ditch network maintenance (DNM) may influence soil water conditions less than initial ditching due to reduced hydraulic conductivity of the peat
- Stand stocking and management substantially influence soil-water conditions
- DNM can lower the GWL and increase tree growth
- DNM growth responses of 0.5–1.8 m3 ha–1 yr–1 during 15–20-years in Scots pine peatland stands reported
- Greatest need for DNM in the early phase of a stand rotation
- Need for better understanding of the link between soil water and tree growth.
Abstract
At sites with either peat or mineral soils in large areas of boreal forests, high soil-water contents hamper tree growth and drainage can significantly increase growth. Hence, areas covering about 15 × 106 ha of northern peatlands and wet mineral soils have been drained for forestry purposes. Usually ditches gradually deteriorate, thus reducing their functionality as drains, and ditch-network maintenance (DNM) might be needed to maintain stand growth rates enabled by the original ditching. This article reviews current knowledge on establishing the need for DNM in boreal forest stands, subsequent growth responses, and the financial outcome of the activity. The issues covered in the review are: (i) ditching, changes in ditches over time and the need for DNM; (ii) interactions between soil water and both stand properties and stand management; (iii) ground-water level (GWL) and tree growth responses to DNM; and (iv) financial viability of DNM. Conclusions about the current understanding of issues related to DNM are drawn and implications for DNM in practice are summarized. Finally, gaps in knowledge are identified and research needs are suggested.
Keywords
peatland;
forest drainage;
conifers;
tree growth;
drainage intensity;
ditch deterioration;
forest water use;
ground-water table;
profitability
-
Sikström,
The Forestry Research Institute of Sweden (Skogforsk), Uppsala Science Park, SE-751 83 Uppsala, Sweden
E-mail
ulf.sikstrom@skogforsk.se
- Hökkä, Natural Resources Institute Finland (Luke), Management and Production of Renewable Resources, P.O. Box 16, FI-96301 Rovaniemi, Finland E-mail hannu.hokka@luke.fi
Received 1 July 2015 Accepted 1 December 2015 Published 17 December 2015
Views 356059
Available at https://doi.org/10.14214/sf.1416 | Download PDF

1 Introduction
In large areas of boreal forests, at sites with either peat or mineral soils high soil-water contents hamper tree growth (Paavilainen and Päivänen 1995; Fenton and Bergeron 2006; Simard et al. 2007). This is because gas exchange between soil and air is much slower in water-saturated soils than in aerated soils (Kozlowski 1982), and consequent limitations in oxygen availability impair functions of plant roots. Hence, a constantly high ground-water level (GWL) leads to shallow rooting, which impairs growth rates, increases mortality and may increase trees’ susceptibility to drought stress during dry spells (Kozlowski 1982; Glenz et al. 2006).
At such sites, lowering the GWL, and thus the soil-water content (SWC) in the unsaturated water-zone, makes conditions more favourable for tree roots. This can substantially increase tree growth, provided that other production factors, e.g. plant-available nutrients, are not growth-limiting. In forestry, SWC are most commonly improved by constructing drainage ditches. In peatlands, drainage is known to improve seedling establishment (Freléchoux et al. 2000) and to increase tree stand density (Hökkä and Laine 1988). In addition, significant tree-growth responses to drainage have been observed in many boreal coniferous forest areas with both peat soils (Seppälä 1969; Payandeh 1973; Hånell 1988; Dang and Lieffers 1989; Lieffers and MacDonald 1990; Gustavsen et al. 1998; MacDonald and Yin 1999) and mineral soils (Trettin 1986; Roy et al. 2000a, Mäkitalo and Hyvönen 2004).
For these reasons, about 15 × 106 ha of northern peatlands, including some areas of wet mineral soils (with no peat or peat-forming vegetation, depending on definitions), have been drained for forestry purposes (Minkkinen et al. 2008). Most of this area is located in Fennoscandia (48%), Russia (28%) and the Baltic countries (11%), and the rest in United Kingdom and Ireland (6%), U.S.A. and Canada (3%) and some odd percent or smaller in Belarus, Poland, Germany and China, respectively, according to the compilation by Minkkinen et al. (2008).
In drained sites gradual deterioration of ditches reduces their water transportation capacity, thereby decreasing their beneficial effects on the ground-water depth (GWD) and SWC (Paavilainen and Päivänen 1995). Therefore, ditch-network maintenance (DNM) might be needed to restore the drainage functions of the ditches to improve tree growth or maintain achieved growth rates (Paavilainen and Päivänen 1995). This can be done by clearing old ditches (ditch cleaning) or by digging new ditches between the old ones (complementary ditching; Ahti and Päivänen 1997). A third possibility is to combine the two operations.
In Finland, a national target has been set to apply DNM in areas of about 8 × 104 ha annually (National... 2011), and during the last two decades it has been applied to between 7 × 104 and 8 × 104 ha–1 yr–1 (Päivänen and Hånell 2012). Most of the area subjected to DNM in Finland is dominated by Scots pine (Pinus sylvestris L.) stands on peatland sites of medium or low productivity. During the last decade, attention has been paid to the need for DNM in Swedish forestry, but no statistics on its extent are available.
In an extensive review on peatland forestry Paavilainen and Päivänen (1995) made some comments on DNM, but knowledge at that time was highly limited and neither standards for the timing of DNM operations nor predicted growth responses could be given. However, some studies had shown that DNM can lower the GWL (e.g. Päivänen and Ahti 1988). In addition, Paavilainen and Päivänen (1995) highlighted the scarcity of studies regarding tree growth responses to DNM. The few published contributions included studies by Stewart (1958) and Olkinuora (1990), which were too short-term (4–5 years) to provide definitive conclusions about DNM’s effects on growth. Paavilainen and Päivänen (1995) also suggested that the drainage capacity is affected by the tree stand present, in addition to the gradient, soil quality, ditching technique and time. Furthermore, they mentioned some qualitative indicators of a need for DNM, including poor tree growth and poor ditch condition. However, they provided no quantitative recommendations, except that complementary ditching should be considered if the original ditch spacing exceeds 50 m (Paavilainen and Päivänen 1995).
According to Hökkä et al. (2008) no easily applicable tools are available for assessing SWC, and the condition of the ditches is often used in practise to evaluate the need for DNM, while other factors that may affect SWC more strongly are ignored. For example, the tree stand itself influences soil-water conditions through evapotranspiration (e.g. Heikurainen 1963; Hökkä et al. 2008). The larger the standing crop (and thus leaf area), the higher the evapotranspiration, which lowers the GWL and SWC. Generally, the natural development of the standing crop results in higher volumes over time and by putting different management operations into practice the development can either accelerate or slow down. Stands’ development and increases in biomass (and hence evapotranspiration) can be accelerated, e.g. by fertilization, or their biomass can be sharply reduced (e.g. by harvesting), thereby affecting the SWC. For example, DNM is sometimes recommended at the time of partial cuttings to compensate for the accompanying reduction in evapotranspiration potential of the tree layer (Ahti and Päivänen 1997; Päivänen and Sarkkola 2000).
After clear-cutting, rises in GWL (Lundin 1979; Päivänen 1980; Dubé and Plamondon 1995) may hamper the survival and growth of the regeneration stocking at some sites (Lieffers and Rothwell 1986; Lieffers and MacDonald 1990; Landhäusser et al. 2003; Jutras et al. 2006). To counteract such rises, construction of new ditches or cleaning of old ditches may be appropriate during the regeneration phase until the ground vegetation and the new tree crop has developed sufficient evapotranspiration potential to maintain desirable soil water conditions without assistance by the ditches (Roy et al. 2000a). Other examples of soil-water regulating operations during the regeneration phase include site preparation and the use of shelterwoods.
At wet sites, the key element of soil preparation is to create a raised spot, a mound, to improve the local soil water conditions for the seedlings (e.g. Sutton 1993; Mäkitalo et al. 2006), while retaining shelterwood trees is likely to buffer GWL rises after harvest in order to maintain suitable soil water conditions for seedling establishment and growth (Hånell 1991; Pothier et al. 2003). Thus, both natural processes and forest management measures influence SWC and the development of forest stands in peatlands and areas with wet mineral soils.
Due to increases in loads of suspended solids (Joensuu et al. 2002; Nieminen et al. 2010), both digging new ditches and DNM may be harmful for downstream surface waters. Therefore, as stressed by Saarinen et al. (1998), it is important to minimize the number of DNM operations from both ecological (impact on aqueous ecosystems) and profitability perspectives. Furthermore, growth responses to DNM may affect global warming potential assessments (Minkkinen et al. 2008), including the ecosystem’s greenhouse gas (CO2, CH4, N2O) emissions and C balances.
In addition to the lack of robust, standard methods for assessing the need for DNM there are uncertainties regarding tree growth responses to it. Some recent studies suggest that DNM does not always affect tree growth in southern and central Finland, where climate is more favourable for evapotranspiration than in northern Finland (e.g. Ahti et al. 2008; Sarkkola et al. 2013). Others have shed some light on stands’ growth responses to DNM (Hökkä 1997; Hökkä and Kojola 2001; Ahti 2005; Ahti et al. 2008), but the findings have not been comprehensively summarized. Thus, there are clear needs for better understanding of appropriate means to establish needs for DNM and tree growth responses to DNM operations.
Thus, this article reviews knowledge regarding establishment of the need for DNM in boreal forest stands, subsequent growth responses and the financial outcome. The review also covers studies on interactions between soil-water conditions and properties of forest soils and tree stands. Based on the findings, principal factors and forest management operations that affect soil water conditions are identified. In addition, requirements for DNM to maintain tree growth cost-effectively, and research needs, are addressed. The review is mainly restricted to studies concerning sites with peat soils (of varying depths), and to a lesser extent drained mineral soil sites. The definition of a peatland site usually includes a minimum thickness for the organic layer, ranging between 20 and 45 cm in some European countries and Canada (Päivänen and Hånell 2012), while sites with shallower organic layers are usually regarded as having “mineral” soils. However, this criterion does not apply in Finland, where a peatland site may be defined in several different ways, for example, dominance (> 75% coverage) by peat-producing genera (primarily Sphagnum) in the ground vegetation (cf. Päivänen and Hånell 2012). The tree species considered in the review are primarily Scots pine and Norway spruce (Picea abies [L.] Karst) . The issues covered are: (i) ditching, changes in ditches over time and needs for DNM; (ii) interactions between soil water and both stand properties and stand management; (iii) responses of GWL and tree growth to DNM; (iv) financial outcomes of DNM; (v) implications for DNM in practice, and (vi) conclusions and research needs regarding these issues.
2 Ditching, changes in ditches over time and needs for DNM
2.1 Ditching aims and drainage intensity
The aim of draining wet peatlands is to increase the air voids in soil, and hence rates of gas exchange. This is because metabolic rates of aerobic organisms (and plant roots) in soil are generally reduced when air-filled porosity falls below 20%, and severely suppressed when it is less than 10% (Glinski and Stepnievski 1985). Thus, these are regarded as minimum thresholds for adequate growth of various tree species, for instance Scots pine in peat (e.g., Paavilainen 1967). In contrast, Wesseling and van Wijk (1957) and Bartholomeus et al. (2008) have shown that several plants can survive and grow in soils in which air content is significantly less than 10%. Nevertheless, numerous studies have shown that drawing down the GWL in peatland sites by ditching can induce increases in peatland tree growth, sometimes remarkably (e.g., Seppälä 1969; Payandeh 1973; Hånell 1988; Frelechoux et al. 2000).
In practice, GWD has been the most frequently used variable in attempts to determine whether air-filled porosity in the rooting zone is sufficient in drained peatland sites. Various authors have proposed target GWDs and ranges when draining various soil types and geographical locations during growing seasons to avoid limiting tree growth. In their synthesis, Vompersky and Sirin (1997) suggest that GWD should ideally range from 30 to 50 cm in poorly decomposed Sphagnum peat, and from 50 to 70 cm in highly decomposed peat. In another synthesis, Paavilainen and Päivänen (1995) recommend GWDs ranging from at least 35 cm in ombrotrophic, undecomposed peat to > 55 cm in well-decomposed, minerotrophic peat. Several authors recommend drainage to maintain the GWL at 20–50 cm between adjacent ditches (Rothwell et al. 1996; Tóth and Gillard 1988; Berry and Jeglum 1991). According to Vompersky and Sirin (1997), Buss and Zalitis (1968) state that the optimal GWD for maximizing growth of pine stands in Latvia on drained peatland is 40 cm, and the soil should not be flooded for at least 30–40% of the growing season. Vompersky and Sirin (1997) also note that Vompersky et al. (1975) concluded that growth of pine in peatlands in northwestern Russia is maximal when the uppermost soil layer is not flooded for at least 85% of the growing season. Similarly, Toth and Gillard (1988) concluded that drained areas should not be flooded for more than 14 consecutive days.
A target GWD range can be attained by using ditches with various combinations of depth and spacing, depending inter alia on the peat depth and wetness, permeability of the mineral soil, gradient, and tree stand properties (Päivänen 1990; Rosen 1989). However, it is very difficult to obtain the desired drainage intensity in every location, using any ditch depth and spacing permutation, due to local variations in soil properties (e.g. peat thickness and water conductivity), stand conditions (e.g. volume and species compositions) and landscape-level variables (e.g. slope and elevation). In practice, 90 cm deep ditches are usually used, at 25–80 m spacing.
2.2 Ditch deterioration
All ditches deteriorate after digging, but at varying rates. Heikurainen (1957) reported that the average depth of hand-dug ditches in peat soils at various sites decreased by 24–36 cm in 20 years, and the most influential factor was peat subsidence, which was positively correlated to the initial thickness of the peat layer. Initial differences in ditch depth diminished due to the deterioration process, leading to the conclusion that a shallow but dense ditch network is optimal for an adequate tree growth rate (Heikurainen 1957; corroborated by Huikari et al. 1966). Timonen (1983) also found that ditches lost a substantial fraction of their depth in a similar timeframe (21–22% in 15 years), with variations largely due to differences in the wetness, thickness and degree of humification of the peat, while the ditching method (using a plow or excavator) had no impact. Half of the ditches Timonen (1983) examined were classified as in need of cleaning after 15 years, mainly due to blockage by vegetation. In addition to subsidence of peat and blockage by vegetation, deterioration of ditches may be caused by collapse of their walls, or obstruction by slash and other materials moved (for instance) by machines being driven across or beside them, erosion and/or silting (Paavilainen and Päivänen 1995).
A logistic regression model for estimating the probability of ditches being in poor condition, using ditch age, geographic location, and peat depth as driving variables, indicated that time elapsed since drainage was the most important factor (Hökkä et al. 2000a). This has been noted in several previous studies (Heikurainen 1957; Timonen 1983; Keltikangas et al. 1986; Isoaho et al. 1993), and can be attributed to the gradual deterioration of ditches with time. In Finland, the probability of ditches being in poor condition 10 and 30 years after drainage was found to be 0.2 and 0.7–1.0, respectively (Hökkä et al. 2000a). Similar probabilities were reached 25–30 years later in southern Finland than in northern Finland. The results also show that deterioration rates are positively correlated with peat thickness (usually measured down to 1 m at the most in many studies), probably due to its association with subsidence of the surface peat (cf. Heikurainen 1957; Timonen 1983). The model constructed by Hökkä et al. (2000a) is used in the MOTTI stand simulator to predict the need for DNM in forecasts of the development of drained peatland stands (Hynynen et al. 2002). All the above mentioned studies suggest that ditches usually deteriorate at a considerable rate from digging onwards. However, following a study of 14- to 26-year-old ditches (on average 50–60 cm deep at the time) in various types of sites, Lauhanen et al. (1998) concluded that ditches’ quality, depth and age provide little indication of the true need for DNM (in accordance with Laine 1986). Thus, they suggested that subjective assessment of ditch status may lead to overestimates of the need for DNM. More generally, subjective visual assessment may provide little indication of either a ditch’s water-transportation capacity or its need for maintenance.
2.3 Tree growth in poorly drained sites
As a ditch network loses its capacity to remove water, the GWL will gradually rise in sites where drainage is mainly dependent on runoff. For example, Sarkkola et al. (2010) showed that ditch depth was inversely correlated with mean late summer GWL in drained peatland sites in Finland.
Inundation of the root layer following rises of the GWL may impair growth of trees. In a field experiment, Pelkonen (1975) observed a significant decline in height growth of Scots pine trees in central Finland (Vilppula Experimental Forest) in the first growing season after lifting the GWL close to the soil surface (< 10 cm) in late July or August in the previous year. Poor condition of ditches or shallow ditch depth was shown as lower tree growth in a study by Heikurainen (1980). Growth models derived from peatland tree growth survey data representing geographically extensive areas also indicate that poor condition of ditches is associated with significant reduction in tree diameter growth (Hånell 1988; Hökkä et al. 1997).
According to Kozlowski (1982) and Glenz et al. (2006) the root inundation parameters that most strongly influence established trees’ functions and growth are the duration and season of the inundation. There are, however, conflicting opinions regarding tree growth responses to seasonal fluctuation of the GWL. According to a review by Glenz et al. (2006), early growing-season flooding is more damaging for plants than late growing-season flooding. However, Pelkonen (1975) (and Päivänen 1984) clearly found that a high GWL had no effect on growth of Scots pine trees in a drained boreal peatland in early summer (before the end of June), but a high GWL in the late growing season (July–August) reduced their growth in the following year. More generally, exposure to flooded (high GWL) conditions is associated with plant injuries, especially during the growing season, when the growing roots have high oxygen requirements (Boggie and Miller 1976; Kozlowski 1982). High GWLs in winter appear to be less harmful for many species (Kozlowski 1982), e.g., Norway spruce (e.g. Wang et al. 2013). Sarkkola et al. (2012) conclude that the GWD should be 35–40 cm in drained peatland during late summer to assure favorable soil-water conditions for tree growth.
2.4 Drainage-induced changes in soils and DNM
Drainage-induced changes generally improve the quality of wet soils as growing media for trees. Lowering the GWL in peatland sites results in increases in substrate aeration (Campbell 1980), soil temperature (Lieffers and Rothwell 1987), rates of decomposition and mineralization (Lähde 1969; Lieffers 1988), and hence availability of plant nutrients (Ponnamperuma 1984). However, Paavilainen (1967) found that increases in average GWD (associated with increases in drainage intensity) resulted in minor increases in the average rooting depth of Scots pine trees. More important factors may be the highest GWL and its duration during the growing season (see section 2.1), because rising GWL rapidly kills roots in the water-saturated zone (Fraser and Gardiner 1967).
Within a few years of first ditching, subsidence of the peat layer (Lukkala 1949; Braekke 1983; Hillman 1992; Rothwell et al. 1996; Minkkinen and Laine 1998a) alters peat’s properties. According to Lukkala (1949) the peat thickness decline by 10–23% and its surface subsides by 7–55 cm on average, these changes being greatest in wet, thick-peated treeless mires and lowest in spruce mires. Lukkala (1949) also found that subsidence was greatest near ditches and occurred during the first five years after ditching. Similarly, Minkkinen and Laine (1998a) concluded that peat in inventoried Finnish pine mires (mean peat thickness, 100 cm) subsided by 22 cm on average in 60 years. They also detected positive correlations between the degree of subsidence and both peat fertility (N concentration) and depth. The main reason for subsidence is the removal of water from the peat pores and subsequent lack of support for the low-density peat soil structure. With time, the weight of the tree stand further increases peat subsidence if tree growth has been strongly promoted.
Increases in aeration of the surface peat after drainage accelerate decomposition, and subsequent humification, thereby increasing the bulk density of the peat. Thus, Minkkinen and Laine (1998b) found that the bulk density of the 80 cm surface peat layer was significantly lower in undrained sites (82 kg m–3) than in sites drained 60 years previously (133 kg m–3). In the 0–20 cm surface peat layer, Silins (1997) observed even faster and larger changes (a 200% increase in bulk density in 7 years). The bulk density of deeper peat layers below the GWL will also increase after drainage, according to Minkkinen and Laine (1998b) and Price et al. (2003). Increases in bulk density decrease peat total porosity and average pore size, which, in turn, reduce soil hydraulic conductivity but increase water retention capacity (Päivänen 1973; Päivänen and Hånell 2012). These observations raise a caveat regarding the assumption that applying DNM to a previously ditched peat site will re-establish soil aeration by lowering the GWL, simply because that is what happened previously, as changes in the soil’s properties will affect the success of DNM. Due to the lower hydraulic conductivity of high bulk density peat, all changes in GWL and SWC will be slower and more difficult to attain than in poorly decomposed peat (e.g., Päivänen 1973). Thus, DNM will have less impact on the ability of gravitational force to drain peat than initial ditching, because in the intervening time the peat has subsidized, resulting in increased bulk density and water retention capacity, and, reduced hydraulic conductivity.
3 Interactions between soil water and stand properties and stand management
3.1 Evapotranspiration of forest stands
Mature tree stands can intercept substantial proportions of gross precipitation during the growing season. For example, Päivänen (1966) reported that Norway spruce, Scots pine and downy birch (Betula pubescens Ehrh.) stands, respectively intercepted proportions of 23–35%, 20–25% and about 20% in drained Finnish peatlands. Similarly, Dubé and Plamondon (1995) found that stands of various mixtures of coniferous and deciduous tree species intercepted 35–41% of the rainfall before harvest at eight sites in Québec, Canada (also Plamondon et al. 1984).
Interception is dependent on several variables, including forest leaf area, stand density and stand height (Aboal et al. 2000). In a Scots pine stand on a drained peatland where the volume of the original tree crop averaged 90 m3 ha–1 Heikurainen and Päivänen (1970) found that thinning at grades of 20, 40, 60, or 100% increased the proportion of precipitation reaching the ground during the first two growing seasons after harvest by 7%, 8%, 12%, and 29%, respectively. During the first growing season after clear cutting the increase was somewhat lower (20%) because the fresh logging residues intercepted some of the precipitation (Päivänen 1980), thus limiting rises of the GWL. This conclusion is corroborated by reports by Dubé and Plamondon (1995) that interception fell to 8–15% after clear-cutting of the stands considered by Plamondon et al. (1984). Heikurainen and Päivänen (1970) also reported that heavier cuttings increased the thickness of the snow layer, and that snow seemed to melt earlier in thinned and clear-cut plots they observed than in untreated plots in drained peatland.
A vigorous tree stand with large leaf area may also significantly decrease soil-water storage under favorable climatic conditions for transpiration (Lagergren et al. 2008; Vincke and Thiry 2008). For example, Heikurainen (1963) found that the GWL clearly falls (sometimes by more than 20 mm) in peat sites supporting such stands on days when rain does not interfere with GWL movements. The magnitude of the changes seems to depend on several variables including the tree species, the stand’s developmental phase, temperature and cloudiness (Heikurainen 1963). Furthermore, Sarkkola et al. (2013) showed that amounts of evapotranspiration and precipitation during the growing-season (May–September) were similar in a drained peatland Scots pine stand in southernmost Finland, but evapotranspiration was substantially lower than precipitation at three more northerly located sites (two of which had lower standing volumes). A very small percentage of precipitation seems to reach the ground through stem-flow in Scots pine and downy birch stands, whereas it seems to be negligible in Norway spruce stands (Päivänen 1966). Plamondon et al. (1984) also found that stem-flow was of minor importance (0.3–3.8% of the rainfall) in four 25 to 65-year-old balsam fir stands.
Sarkkola et al. (2010) conclude that the effect of stand volume on the GWL during “normal” rainy summers in peatlands is probably due to high canopy interception in well-stocked stands, since interception reportedly influences their water balance more than transpiration (Heikurainen 1963; Laine 1984; Dubé et al. 1995). Dube and Plamondon (1995) also conclude that the reduction of interception is the most important contributor to the common post-harvest rise of the GWL. Furthermore, Dube and Plamondon (1995), and Päivänen (1980), recommend that regeneration stocking and other vegetation, e.g. shrubs and the field-layer, should be protected and logging debris left on site after cutting in order to maximize interception and evapotranspiration. Thus, stand stocking and management substantially influence interception, evapotranspiration and hence soil-water conditions.
3.2 Stand stocking, stand development and GWL
Several studies have quantified correlations between stand stocking or stand growth and the GWL. The volume of the tree stand is the most influential parameter for the GWD according to Laine (1986), explaining more than 50% of the GWD variation in drained peatlands. A 10 m3 ha–1 increase in stand volume was associated with a 2 cm lowering of the GWD (Laine 1986), while Ahti and Hökkä (2006) found that increases in growth rate of 1 m3 ha–1 yr–1 or stem volume of 10–15 m3 ha–1 were associated with falls in the GWL of ca. 1 cm in late summer and autumn, but not in spring or early summer. Sarkkola et al. (2010) report a similar relationship, a 10 m3 ha–1 increase in stem volume corresponding to a 1 cm lower GWL during the growing season, while Heikurainen (1980) found that an increase in stand volume of about 15 m3 ha–1 or volume increment of about 0.6–0.7 m3 ha–1 yr–1 corresponded to up to a 10 cm lowering of the GWL. All of these reported correlations may potentially be valid up to a certain stand volume, as suggested by findings that at volumes exceeding 150–200 m3 ha–1 the GWL may have little dependence on stand volume (Sarkkola et al. 2010). Local, within-stand spatial variation in stand stocking has also been shown to correlate with local variation of GWD (Haahti et al. 2012).
Hökkä et al. (2008) have developed a model to quantify the dynamic interactions among stand volume, volume growth, and GWL, illustrating the interrelationship between GWL and tree-stand growth. According to the model, a deep initial GWL results in high stand growth, and a further increase in stand volume subsequently reduces the GWL. The model can be used for simulating stand volume development over 20 years in southern boreal drained peatlands supporting Scots pine forests. It can also predict the GWL under given stand conditions and management practices (Hökkä et al. 2008).
Fertilization promotes tree growth in most boreal forests in both peatlands (e.g. Hökkä et al. 2012) and upland sites with mineral soils (e.g. Kukkola and Saramäki 1983; Pettersson 1994). It may also have rapid, strong, and favorable effects on soil-water conditions, due to increases in evapotranspiration mediated by associated increases in leaf area. In a Scots pine experiment reported by Heikurainen and Päivänen (1970) GWL was lowered by 5–8 cm and the runoff by c. 25 % during the second vegetation period after fertilization with phosphorus (P) and potassium (K). Observed increases in needle length and needle mass suggested that interception had increased, and consequently throughfall had decreased by about 5%. The cited authors concluded that fertilization increases evapotranspiration, and that the hydrological effects are both rapid and substantial. Päivänen (1980) also observed a fall in GWL (of 1–4 cm) after fertilization of mature Scots pine and Norway spruce stands (with 110–120 and 140–200 m3 ha–1 stocking, respectively). Furthermore, Ernfors et al. (2010) found that GWL was significantly lower during the fifth growing season after applying wood ash in a Scots pine stand on a drained oligotrophic bog in southern Sweden. During the first five years after the application the basal area increment increased by 20%.
3.3 Biomass removal and GWL
Several studies have investigated effects of removing biomass on soil-water parameters, especially the GWL (Table 1). After timber harvesting, GWL rises have been observed in both boreal forested peatlands (e.g. Heikurainen 1967; Dubé et al. 1995; Roy et al. 1997; Roy et al. 2000b, Lundin 2000), and mineral soil sites (e.g. Lundin 1979; Dubé and Plamondon 1995). In recently harvested sites, drainage is known to reduce GWL rises (e.g. Ahti and Päivänen 1997; Päivänen and Sarkkola 2000), thereby limiting further paludification and improving rooting-zone conditions (Roy et al. 2000a) for seedling growth.
| Table 1. Biomass removal and changes in ground water level (GWL) recorded in indicated studies. | |||
| Biomass removal 1 | Δ GWL 2 (cm) | Note 3 | Reference |
| Partial removal, peat soils | |||
| PCT (from 3.5–10.4 m2 ha–1 to 0.4–2.5 m2 ha–1) | c. 2 | Mean rise from June to August, during 2 years monitored. A.b. and L.l. dominated stands at 4 sites in Quebec, Canada (organic top layer > 50 cm). | Jutras et al. 2006 |
| TBA 20–50% | 1.7 – 2.3 cm. | P.s. on 4 drained peat (depth > 90 cm) sites in N Finland. IS 70–110 m3 ha–1. | Hökkä and Penttilä 1995 |
| TVO 10% TVO 15%, 28% | ± 0 ≤ 7 | P.s. on a drained bog in S Finland. IS = c. 95 m3 ha–1. | Päivänen and Sarkkola 2000 |
| TBA 0%, 20%, 30%, 50% | c. 2–5 (data from figure). GWL lowest in 0% and highest in 50%; non-significant differences. | Drained P.a. mire in E Finland. IS 200–350 m3 ha–1. DC concurrently with T. | Hökkä et al. 2000b |
| T 17%, T 26% | 0–15 The heavier cutting the larger the GWL change. | Mature P.s. and P.a. stands on drained peat in S Finland with volumes of 107–120 m3 ha–1 (P.s.) and 137–198 m3 ha–1 (P.a.). | Päivänen 1982 |
| T 17% T 30% | 0–9 5–15 | P.s. (IS = 110–120 m3 ha–1) N.s. (IS = 140–200 m3 ha–1) in drained peatlands. | Päivänen 1980 |
| T 20–50% | A few cm, but clearly discernible. | Seven exp. in P.s. and B.spp. in drained peatlands in central Finland. IS = 50–150 m3 ha–1. | Heikurainen 1967 |
| T 20%, 40% T 60% | 0–4 5–7 | P.s. in a drained peatland in central Finland. IS = c. 90 m3 ha–1. | Heikurainen and Päivänen 1970 |
| T (SC) | 2–13 | Four P.a. stands on mires in central Sweden. Mean of three years after harvest. | Lundin 2000 |
| Partial removal, mineral soils | |||
| PCT (from 3.5–10.4 m2 ha–1 to 0.4–2.5 m2 ha–1) | 1.5–3 | Mean rise from June to August, during 2 yrs monitored. A.b. and L.l. dominated stands at 2 sites in Quebec, Canada (organic top layer < 20 cm). | Jutras et al. 2006 |
| T 0%, 40%, 50%, 60% | GWL rise linearly related to cutting percentage. | P.r. and A.b. dominated stand on gleyed humoferric podzol (organic top layer c. 10 cm) in Quebec, Canada. IS = 40 m3 ha–1. | Pothier et al. 2003 |
| Final cutting, peat soils | |||
| FC | 4 | Organic soil. | Marcotte 2005 (in Jutras et al. 2006) |
| FC | 2–27 | P.s. (IS = 110–120 m3 ha–1) and P.a. (IS = 140–200 m3 ha–1). | Päivänen 1980 |
| FC | 20–40 | Seven exp. in P.s. and B.spp. in drained peatlands in central Finland. IS = 50–150 m3 ha–1. | Heikurainen 1967 |
| FC | 5–14 cm | P.s. in a drained peatland in central Finland. IS = c. 90 m3 ha–1. | Heikurainen and Päivänen 1970 |
| FC | c. 22 cm | P.r. and A.b. dominated stand on gleyed humoferric podzol (organic top layer c. 10 cm) in Quebec, Canada. IS = 40 m3 ha–1. | Pothier et al. 2003 |
| FC | No significant GWL rise after shelterwood removal relative to some years after shelterwood cut. | FC of shelterwoods, treatments 40%, 50%, 60% in the study by Pothier et al. (2003) See above. | Prévost and Gauthier 2013 |
| FC | 4–11 | Four P.a. stands on peatlands in central Sweden. Mean of three years after harvest. | Lundin 2000 |
| FC | 18 (12–37 within site variation) | P.s. on a drained peatland in S Finland. IS = 100 m3 ha–1. | Sarkkola et al. 2013 |
| FC | 17 | P.a. on peatland in Sweden. IS = 300–400 m3 ha–1. | Lundin 1999 |
| FC | No GWL rise at 20 m ditch distance. At 40 and 60m ditch distances GWL rise occurred 10–30 m away from the ditches. | P.m. dominated stand on a fen with highly humified peat as top soil (100 cm) in Quebec, Canada. IS = 60 m3 ha–1. | Jutras and Plamondon 2005 |
| FC | 4–7 | Five fens, bogs and swamps (peat thickness > 30 cm). Study in Quebec, Canada, in a flat lowland forested wetland area with poor drainage. A variety of tree species included, e.g. A.b., P.r., P.m., A.r. and T.o. | Dubé et al. 1995; Dubé and Plamondon 1995 |
| FC | 12–16 cm, depending on total removed volume | Mature P.a. and P.s. stands in southern Finland, volumes removed 138 m3 ha–1 and 114 m3 ha–1. | Päivänen 1982 |
| Final cutting, mineral soils | |||
| FC | 10 | Mineral soil. | Marcotte 2005 (in Jutras et al. 2006) |
| FC | 20 | Two gleyed mineral soil sites (thickness of organic layer 10–30 cm). Study in Quebec, Canada, in a flat lowland forested wetland area. A.b. and P.r. dominated stands. | Dubé et al. 1995; Dubé and Plamondon 1995 |
| FC | 31 | Mean during 4 years after harvest. P.s. and P.a. dominated stands (IS = 100–200 m3 ha–1) on upland morainic mineral soil in central Sweden. | Lundin 1979 |
| 1 PCT = Pre-commercial thinning; T = Thinning; TBA = Thinning by stem basal area; TVO = Thinning by stem volume; SC = Shelterwood cut; FC = Final cutting. | |||
| 2 Commonly given as mean change in ground water level (GWL) over the vegetation period. | |||
| 3 IS = Initial stocking before harvest; DC = Ditch cleaning; A.b. = Abies balsamea (L.) Mill.; L.l. = Larix laricina (Du Roi) K. Koch; P.s. = Pinus sylvestris L.; P.a. = Picea abies (L.) H. Karst.; P.r. = Picea rubens Sarg.; P.m. = Picea mariana (Mill.) Britton, Sterns & Poggenb.; B.spp. = Betula spp.; A.r. = Acer rubrum L.; T.o. = Thuja occidentalis L. | |||
Jutras et al. (2006) found that pre-commercial thinning to c. 2300 stems ha–1 (from a basal area of 3.5–10.4 m2 ha–1 to 0.4–2.5 m2 ha–1 caused an average GWL rise of 3 ± 5 cm (mean ± SD) between early June and the end of August, during two monitored years at six experimental sites in Quebec, Canada (Table 1). The rise of the GWL was correlated with the basal area removal, although an increase in removal from c. 0.1 to 4 m2 ha–1 only resulted in a 2 cm rise in the GWL (from 4 to 6 cm). After the previous harvesting in 1991, 12 years before the focal pre-commercial thinning, the GWL had risen by 10 cm on average in the mineral soil sites and 4 cm on average in the peatland sites (Jutras et al. 2006).
However, Hökkä and Penttilä (1995) found that a thinning grade of 20–50% caused at most a slight rise of the GWL in Scots pine stands on drained peatlands in northern Finland (Table 1), possibly because the GWL was initially high. They suggested that the stand may have little influence on the GWL in northern Finland because of low growth and transpiration rates, and the large volume of water stored in the soil due to the humid climate. In addition, the ditches were cleaned a few years before the thinning at three of the four studied sites.
Numerous other authors have addressed changes in the GWL following thinning or clear-cutting (with or without other practices) (Table 1). Reported observations include the following. Päivänen and Sarkkola (2000) found that thinning of 15% and 28% (by stem volume) in southern Finland raised the GWL by about 7 cm at most. Subsequent DNM restored the GWL to the pre-thinning level, and 10% thinning had no influence on the GWL. Hökkä et al. (2000b) studied GWL rises after thinning with 0%–50% grades in a drained Norway spruce stand on peat soil in eastern Finland. Generally, the GWL was lowest in the non-thinned plots and highest in the 50%-thinned plots, but the between-treatment differences were not statistically significant. The cited authors attributed the absence of any thinning effect to a concurrent ditch cleaning operation. A constructed regression model indicated that the pre-thinning GWL, volume removed, standing volume and local basal area (within 3 m around a measurement tube) significantly influenced the GWL at the measurement points (Hökkä et al. 2000b). (Päivänen 1982) found that thinning at grades of 17% and 26% induced rises of 3–16 cm in the GWL in Scots pine- and Norway spruce-dominated stands in a drained peatland in southern Finland. In addition, Päivänen (1980) detected 0–9 cm rises in the GWL after thinning (at 17% grade) in mature stands of Scots pine and Norway spruce, greater rises after ca. 30% thinning, and even greater rises after clear-cutting (Table 1). Päivänen (1980) suggested that the variation in the magnitude of the change was partly due to variation in peat properties. Heikurainen (1967) also found that the GWL rose in seven experimental mature Scots pine and downy birch stands in central Finland, by 20–40 cm, during two monitored years after clear-cutting. The change in the GWL was positively correlated with the depth of the pre-treatment GWL, and 20–50% thinning had a weaker, but clearly discernible effect (a few cm).
Heikurainen and Päivänen (1970) also found effects of both thinning and clear-cutting on the GWL in a Scots pine stand on a mire in central Finland. At GWLs of 25–55 cm in the control plot the GWL rose 0–4 cm after 20% or 40% thinning, 5–7 cm after 60% thinning, and 5–14 cm after clear-cutting (Table 1). The cited authors concluded that the observed changes in GWLs were rather small compared with previously recorded changes, for instance those reported by Heikurainen (1967), possibly because ditches in the sites provided good drainage.
Similar responses have also been recorded elsewhere. For example, Pothier et al. (2003) observed GWL rises during the year following cuts, which were linearly related to basal area removal (0–100%), in a stand dominated by red spruce (Picea rubens Sarg.) and balsam fir (Abies balsamea [L.] Mill) on a lowland mineral-soil site in eastern Canada (Table 1). During subsequent years, the GWL in the treated plots gradually approached the pre-treatment level and after five years there was no clear correlation between the thinning grade and GWL. However, the GWL was still higher in the clear-cut plots than in control plots. The cited authors suggested that the GWL recovery was related to increases in leaf biomass of the regeneration stratum over time, rather than crown expansion of residual trees. The final removal of the shelterwoods (thinning grades 40%, 50% and 60%) did not lead to any significant GWL rise during the following five years relative to the GWLs a few years after the initial shelterwood cut and clear-cut (Prévost and Gauthier 2013). This was most likely due to the development of a new cohort, which contributed to increased evapotranspiration and counteracted any potential rise in GWL. The GWLs had not fully recovered to pre-treatment levels in the original stand (control 0%).
Lundin (2000) observed corresponding rises in GWL after shelterwood cutting and clear-cutting of Norway spruce stands on peat soils at four sites in Central Sweden. A greater GWL rise, of 17 cm, was observed during the four years after clear-cutting of a Norway spruce stand in a drained spruce mire in southern Sweden with 0.2–2 m thick peat (Lundin 1999). Sarkkola et al. (2013) recorded a similar GWL rise after clear-cutting of a Scots pine stand on a drained peatland of dwarf-shrub site type in southern Finland, with 100 m ditch spacing and more than 1 m thick peat.
Jutras and Plamondon (2005) studied the effects of clear-cutting on the GWL at five distances from ditches (3, 6, 10, 20 and 30 m) with three ditch spacings (20, 30 and 40 m) in an old Canadian stagnant black spruce-dominated forest on a site with highly humified fen peat as top soil overlaying a fine-textured mineral soil. The site was drained 10 years before harvesting. After the drainage, the GWL was lowest at 3 m from the ditches, at each of the ditch spacings, and the lowering of the water table at 6, 10 and 20 m from the ditches decreased as the spacing increased. The high degree of decomposition of the peat below 10 cm – classified as 7 and 10 according to von Post at 10 and 60 cm depths, respectively – was connected to low hydraulic conductivity, which probably contributed to the smaller lowering of the GWL away from the ditches. After harvest no rise in GWL was observed in plots with ditches at 3–20 m spacing, and with wider spacing no rise at 3 m from the ditches was recorded, but rises were registered at greater distances (10, 20 and 30 m). Jutras and Plamondon (2005) concluded that the GWL rose when drainage before harvest did not lower it by more than 10 cm.
Dubé et al. (1995) and Dubé and Plamondon (1995) respectively recorded average GWL rises of 20 cm after clear-cutting of two stands on gleyed mineral soils, and 4–7 cm in five stands growing on peaty gley soils in three fens and two bogs, in a flat forested wetland area with poor drainage in Quebec, Canada. Thus, the rises were generally smaller in bogs and fens. They also found that the lower the pre-cutting GWL, the larger the rise after cutting. In the wettest site, where the pre-treatment GWL was < 10 cm from the ground surface, the GWL dropped by 3 cm after clear-cutting, probably due to evapotranspiration from exposed water surfaces. Dubé et al. (1995) suggest that the transitional sites, i.e., sites between the bogs or fens and the uplands, were most susceptible to hydrological changes after final cutting. They also conclude that clear-cutting using narrow strips is not an effective solution to prevent water table rises, and suggest a more efficient way would be to leave logging debris and, if present, small trees and advance growth, in order to maintain interception and transpiration.
In a follow-up study, Marcotte et al. (2008) monitored the GWL recovery after clear-cutting and drainage. Ten years after clear-cutting, the GWL in undrained plots was still 5–7 cm higher than the pre-cut levels. The slight recovery of the GWL in these plots registered in the first years after clear-cutting leveled off after the third year. Based on this and another study in the region (Pothier et al. 2003), Marcotte et al. (2008) suggest that full GWL recovery after clear-cutting will take more than 6–10 years. Ten years after cutting, the rainfall interception by vegetation had reached nearly 50% of the pre-cut levels, indicating that both the present vegetation and subsequently regenerating vegetation play important roles in the water balance. However, complete recovery may need a more densely standing crop and eventually a more complete occupation of the soil by deeper tree roots as suggested by Marcotte et al. (2008). In the same study, the immediate GWL drawdown following drainage mitigated the GWL rise induced by clear cutting up to 40 m from ditches on mineral soils. On peat soils, the GWL was significantly lowered by the ditching up to 20 m into the uncut forest and up to 60 m into the clear-cut area.
It should be noted that Lundin (1979) also observed very strong effects of clear-cutting on GWL in upland morainic mineral soils in central Sweden; a 31 cm rise in mean GWL (from the original 92 cm below the ground surface) during the following four years. However, in contrast to many of the findings cited above, Mannerkoski et al. (2005) did not detect any significant change in the GWL in two catchments where 10% or 30% of the area was clear-cut, disc-ploughed and planted with Scots pine and Norway spruce seedlings, then monitored during the five years following the harvest.
In summary, stocking clearly reflects the capacity of a forest stand to intercept rainfall and take up soil-water, at least for a vigorously growing stand (e.g., Le Maitre and Versfeld 1997; Hökkä et al. 2008). Furthermore, as stated by Sarkkola et al. (2010), it is well established that evapotranspiration of forest vegetation, mainly of the forest stand, directly affects the GWL, not only in sites with drained peat soils (e.g. Heikurainen 1963; Ahti 1978; Ahti and Hökkä 2006; Hökkä et al. 2008) but also in mineral soil sites (e.g. Lagergren et al. 2008; Vincke and Thiry 2008). In boreal peatlands, this relationship seems to apply across wide ranges of site and climatic conditions (Sarkkola et al. 2010). Accordingly, an increase in stand biomass lowers the GWL, while cuttings have the opposite effect.
4 Responses of GWL and tree growth to DNM
Both deterioration of ditches (section 2.2) and biomass removal (section 3.3) may lead to rises of the GWL that hamper tree growth, implying a need for DNM. The greatest GWL rises in drained areas are usually caused by radical cuttings (Table 1).
Ahti and Päivänen (1997) reported responses of GWL and tree growth to DNM recorded in experiments at 12 sites with Scots pine stands (20–80 m3 ha–1; Päivänen and Ahti 1988) in drained peatlands. After cleaning the ditches to their original depth, digging new ditches between the old ones, and combining these treatments, the GWL was lowered by 4, 6 and 10 cm on average, indicating that they have additive effects (see also Päivänen and Ahti 1988). The lowering of the GWL was statistically significant in five (of eight) experiments (Ahti and Päivänen 1997). A similar lowering of the GWL after ditch cleaning was also reported by Päivänen and Sarkkola (2000). In addition, Koivusalo et al. (2008) found a significant, much greater (30–40 cm) lowering of the GWL after ditch cleaning in experimental sites in northern Finland with a thin peat layer (12–33 cm) and sandy subsoil, but no significant change in GWL after cleaning in sites with deeper peat.
In Ahti and Päivänen (1997) indications of increased tree growth following DNM were observed in half of the 12 experiments, but the growth analysis was limited by the short monitoring period (5–8 years) following DNM, and no statistical analyses of the growth data were conducted. However, in many of the experiments showing increased growth, particularly those located in the north, complementary ditching appeared to promote growth considerably more than ditch cleaning, and the growth responses occurred mainly in trees close to the new ditches (Ahti and Päivänen 1997).
A later analysis, by Lauhanen and Ahti (2001), of the experimental material examined by Ahti and Päivänen (1997), showed that ditch cleaning, complementary ditching and the combination of both had increased volume growth by 0.16 (not statistically significant), 0.36 and 0.48 m3 ha–1 yr–1, respectively, after 10 years. Furthermore, regardless of the initial stand volume, increased growth was detected during the second five-year period, and the average growth responses were considerably higher during the second five-year period than during the first period (Lauhanen and Ahti 2001). Ten years after DNM, both the cleaned and complementary ditches were classified as being in a better condition and significantly deeper (78 and 73 cm, respectively) than non-maintained control ditches (41 cm deep). At most sites the drainage was considered sufficient according to the authors. However, no dramatic reductions in stand growth were detected in untreated control plots during the 10-year post-treatment period, which raises questions about both the timing and need for the DNM operations applied in the monitored stands.
In a further subsequent study of the experimental material used by Ahti and Päivänen (1997) and Lauhanen and Ahti (2001), Sarkkola et al. (2012) found that the 20-year growth response, in terms of increases in average annual volume growth and relative volume growth, were negatively related to the GWL before the DNM. The largest growth responses to DNM were observed in Scots pine stands where the pre-treatment GWD was 25–30 cm or shallower, while in stands where it was deeper than 35–40 cm there were minor growth increases. Sarkkola et al. (2012) also observed high variability in growth responses to DNM in stands where the pre-treatment GWD was 20–35 cm, concluding that other factors also strongly influence the optimal timing for DNM to obtain adequate growth responses. The growth responses reported by Sarkkola et al. (2012) averaged 0.6 m3 ha–1 yr–1 at sites in northern Finland, and 1.1 m3 ha–1 yr–1 at sites in southern Finland, during the 20-year effect period. In this study, DNM included ditch cleaning and complementary ditching combined.
Sarkkola et al. (2010) concluded that in stands with volumes exceeding about 150 m3 ha–1, the condition of the ditches had a marginal influence on stand development, suggesting a weak or non-existent growth response to DNM. This conclusion was corroborated by the finding that evapotranspiration amounts were similar to growing-season precipitation in a Scots pine stand on a drained peatland in southernmost Finland, suggesting that there was no need for DNM (Sarkkola et al. 2013). However, as Sarkkola et al. (2010) noted, it is not possible to identify a need for DNM solely from stocking criteria, as weather conditions and peat properties also affect the GWL.
Lauhanen et al. (1998) evaluated effects of complementary ditching using data obtained from surveys of permanent growth sample plots on peatlands in northern Finland. In “pine mires”, the stand growth was 0.6–1.0 m3 ha–1 yr–1 higher over about 20 years with complementary ditching than solely with initial ditching. In “spruce-birch swamps” the growth was also higher in stands where complementary ditching was conducted (by 1.5–1.9 m3 ha–1 yr–1 over about 20 years) compared to stands with initial ditching only.
Simulations by Hökkä (1997), based on the experimental data published by Ahti and Päivänen (1997), indicated that DNM induced 0.5–1.8 m3 ha–1 yr–1 increases in annual growth during the following 15 years. The smallest responses were found in northern Finland following ditch cleaning. Using a large dataset, combining data acquired from observations of experimental sites and routinely surveyed stands, with and without DNM treatment, Hökkä and Kojola (2003) constructed a model for predicting growth responses to DNM. The model predicted that average responses to DNM (ditch cleaning, complementary ditching and a combination of the two pooled together) in northern Finland would be growth increases of 0.50–0.86 m3 ha–1 yr–1 during a 15-year simulation period in two example stands. The driving variables were stand basal area, peat thickness, temperature sum and site type (classified as originally forested sites or poorly stocked wet sites) (Hökkä and Kojola 2003). Similarly, from an analysis of experimental stands, Ahti (2005) concluded that DNM in peatland stands is likely to induce growth increases of 0.4–0.7 m3 ha–1 yr–1 over 15 years. Finally, based on data from the same experiments, Ahti et al. (2008) showed that over a 20-year study period DNM increased growth in northern Finland (0.6–1.1 m3 ha–1 yr–1) but had almost negligible effects in southern and central Finland (0–0.4 m3 ha–1 yr–1), especially if ditch cleaning was the method applied. In northern Finland the response to ditch cleaning, complementary ditching and combination of both was more significant and also lasted longer, i.e., at least 20 years (Ahti et al. 2008).
Lauhanen and Ahti (2001) suggest that there were no growth increases following DNM in some monitored stands, especially pine stands in southern Finland, because there was no acute need for DNM, even if the ditches were in poor condition. As the stands developed, the accompanying increases in evapotranspiration (see section 3) probably compensated for the reduction in drainage capacity of the deteriorating ditch system. In such situations the GWL will remain deep enough to sustain appropriate tree growth as long as the evapotranspiration of the stand remains sufficiently high, and the shallow ditches will still be deep enough to collect the surface runoff. Furthermore, the rather limited growth responses of the pine stands indicate that the hydrological need for DNM was less urgent than expected. Lauhanen and Ahti (2001) also concluded that even if the maintenance of the ditch system is necessary, delays in performing it will probably not cause dramatic growth reductions or irreversible changes in the development of the stands.
When attempting to quantify growth responses of peatland stands to DNM, a significant problem is identifying an appropriate reference growth rate (Hökkä and Kojola 2003). Since a gradual rise in the GWL induces a slow gradual decline in growth, the response to DNM is also dependent on the degree of growth limitation at the time of treatment, as noted by Sarkkola et al. (2012). See also Fig. 1. In most cases when DNM is implemented growth will probably have been sub-optimal for some time, but even if growth is close to optimal there may still be a positive response if the stand’s growth is likely to decline soon without treatment (Ahti and Päivänen 1997). In research, the reference growth problem has been solved in two ways. Data obtained from observations of experimentally treated material are usually compared to corresponding data for non-treated controls (Ahti and Päivänen 1997; Hökkä 1997; Lauhanen and Ahti 2001; Ahti 2005; Ahti et al. 2008; Sarkkola et al. 2012). A drawback of this approach is the possibility of bias related to incomplete isolation of treatments in the experimental lay-out (e.g. Koivusalo et al. 2008). The other commonly applied approach (notably when using increment boring survey data) is to compare growth rates before and after DNM (Hökkä and Kojola 2001), which may lead to underestimates of the response because it does not account for the possible decline in growth of non-treated stands. Thus, Hökkä and Kojola (2003) combined data from several sources to obtain a generalizable estimate of the growth response. However, in all situations the responses will vary, depending on the soil water conditions when DNM is implemented (Fig. 1).
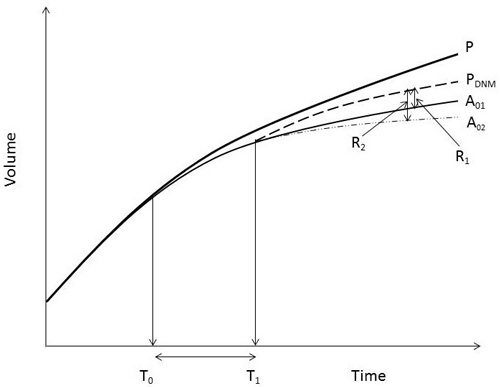
Fig. 1. A schematic figure on stand stem volume development under different assumed site water conditions. P = potential volume development if there is no need for ditch network maintenance (DNM) operation. A01, A02 = alternative development after time point T0, when soil water conditions begin to limit growth moderately (A01) or severely (A02). T1-T0 is the period of sub-optimal growth for senarios A01 and A02. PDNM = potential development if DNM operation is carried out at time point T1. R1 and R2 represent growth reponses of DNM in relation to the two senarios with growth limitation without DNM.
5 Financial outcome of DNM
To be a cost-effective management operation, DNM must either increase growth of stands or prevent decline of their growth in the future. Whether or not the operation is profitable will depend on the magnitude of growth increases, investment costs, interest rates, timber prices and pay-back time. However, it should be kept in mind that DNM is commonly part of a management regime, encompassing operations from regeneration to final harvest and may also include one or two thinnings. Thus, the economical outcome of a separately conducted DNM may not be the same as the outcome of DNM simultaneous with thinning (cf. Fig. 2) as a part of an overall management regime (cf. Ahtikoski et al. 2008; Kojola et al. 2011).
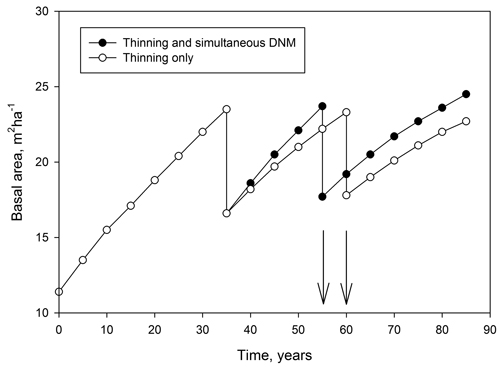
Fig. 2. An example of the impact on basal area development of ditch network maintenance (DNM) operation for a management schedule of a peatland stand when DMN is conducted at the time of first commercial thinning (basal area 12 m2 ha–1 at the start of the simulation). Without DNM the second thinning will be done five years later (see arrows) due to the lower growth rate of trees. Earlier incomes from thinning generally improve the profitability of the schedule including DNM. Data generated by the simulation program MOTTI (Hynynen et al. 2002).
In addition, to calculate the economic outcome of a DNM operation empirical data covering the whole growth–response period are required. Alternatively, yield data generated by simulations can be used. For example, the MOTTI stand simulator (Hynynen et al. 2002) includes a model to predict the growth response to DNM (Fig. 2) (Hökkä and Kojola 2003). However, if simulations are used the accuracy of the results will inevitably depend on the accuracy of the model used to simulate the timber yields (Ahtikoski et al. 2012).
Aarnio et al. (1997) calculated an internal rate of return on investment of 0–9% for DNM performed in four experimental stands, based on the growth response over 10 years (using the same data as Ahti and Päivänen 1997). Profitability was found to be low for both the poorest, low-stocked sites and productive sites with high initial stand volume, while it was highest for medium-productivity fully-stocked stands, in which ditches required maintenance (Aarnio et al. 1997).
According to simulations by Hytönen and Aarnio (1998) DNM can be profitable in terms of both net-present value and internal rate of return, at least if the DNM involves complementary ditching or a combination of complementary ditching and ditch cleaning. Ditch cleaning alone generally yielded negative net present values at interest rates of 3% and 5%. In terms of internal rate of return, ditch cleaning yielded 2.5%, complementary ditching 6.8% and the combination of the two 5.5%. Thus, complementary ditching was more profitable than ditch cleaning. Only the DNM costs were included in the calculations, which were based on data from nine drained pine-dominated oligotrophic peatlands (some of the sites reported by Ahti and Päivänen 1997 and Lauhanen and Ahti 2001). The benefit due to DNM was determined as the difference in value of growing stock between treated plots and untreated control plots.
Ahtikoski et al. (2008) also analyzed the profitability of a DNM in terms of return of investment and net-present value. They used the MOTTI stand simulator (Hynynen et al. 2002) to forecast development of 44 randomly selected normally-managed drained peatland stands in northern Finland with or without DNM and calculated their long- and short-term financial performance. At a given cost, DNM was most profitable in stands with high initial stocking, growing on fertile types of sites with a high temperature sum. The initial stocking appeared to have most influence on the financial performance. The minimum profitability threshold seemed to be relatively low in terms of stand basal area (4–6 m2 ha–1). Another conclusion was that if the monetary value of the accumulated growth cannot be realized until the final cutting, the profitability of DNM remains marginal. This reflects the impact of initial stocking. Densely stocked stands can be commercially thinned, enabling harvest income to be generated at an earlier stage from the additional yield generated by DNM. The mean annual increments without DNM over the simulated 45-year period ranged between 1.9 and 4.0 m3 ha–1 yr–1 for the two stand and site types analyzed. DNM increased the mean annual increments by about 10% (0.24 m3 ha–1 yr–1 and 0.42 m3 ha–1 yr–1, respectively). The return of investment was between 1.8% and 3.0%. The net present value with and without DNM was compared for the whole rotation period, and found to be 4–14% higher for the alternatives with DNM than for those without it. The results are applicable to forest stands on peatland sites that have been managed according to standard silvicultural recommendations and subjected to initial ditching.
Ahtikoski et al. (2012) conducted a study to determine the optimal management regime in terms of net present value for 17 drained peatland stands in northern Finland. Possible management variables included timing of ditch network maintenance, commercial thinning and final cutting as well as intensity of thinning (thinning grade), as recommended by the current guidelines. Higher interest rates led to shorter rotations and less intensive management schedules. They also found that in the harsh northern climatic conditions lower mean annual increment resulting from extensive management could result in a higher net present value. Consequently, DNM may be an unprofitable operation, especially in low-stocked northern stands, even if ambient drainage conditions are poor (Ahtikoski et al. 2012).
6 Implications for DNM in practice
6.1 Operational implications
The primary aim of DNM in established forest stands is to sustain tree growth in ditched sites. As concluded by Lauhanen et al. (1998), Laine (1986), and Sarkkola et al. (2012), visual inspection of the status of ditches may lead to overestimates of the need for DNM. The lack of a convenient, robust method for visually classifying the water transportation capacity of a ditch in the field, and its influence on tree growth, makes it difficult to judge the need for, and optimal timing of, DNM. However, the time since the latest ditching operation is a potential indicator (section 2.2). For example, the probability that DNM will be needed is quite high after 25–30 years, and very high after 50 years (Hökkä et al. 2000a).
It is well established that the present forest stand strongly influences the water balance at a forested site and, consequently, the GWL and SWC (section 3). In studies cited in section 3.1 proportions of precipitation intercepted varied between 20% and 40% (in stands with stem volumes of about 100–200 m3 ha–1). Thus, canopy interception (and evapotranspiration) significantly affect the soil-water conditions. Accordingly, partial (thinning) or total removal (clear-cutting) of the tree biomass usually raises the GWL. The studies cited in section 3.3 suggest that GWL generally rises by 0–15 cm after thinning and up to 40 cm after clear-cutting. The magnitude of the rise is dependent on, (i) the pre-treatment GWL; the deeper initial level the greater rise, (ii) the size of the biomass removal; the greater removal the greater rise, and (iii) the hydraulic conductivity of the soil; the lower conductivity the greater rise of the GWL. In the few reported studies of effects of DNM in sites with mineral soils, the GWL rise was generally larger than in peat soils, probably because both the initial GWL and the hydraulic conductivity of the soils were lower. If stand properties are also considered, the need for DNM is greatest in the early phase of a stand rotation period, especially after clear-cutting, which usually induces a rise in the GWL when a new tree generation is about to be established. A need for DNM may also arise after partial early cuttings (first commercial thinning), but at later phases in more highly stocked stands with volumes exceeding 150 m3 ha–1 a weak growth response to a DNM operation is suggested based on results of Sarkkola et al. (2010) concerning the minor impact of ditch condition on GWD in such stands. Hence, the costs and potential benefits of DNM in highly stocked stands must be carefully evaluated. Growth responses to DNM may also be weak if the pre-treatment GWL is deeper than 35–40 cm (Sarkkola et al. 2012).
Regarding site conditions, the texture of the soil must be taken into account. Well-humified peat (with high bulk density) and fine-textured mineral soils have high water-retention capacity and low hydraulic conductivity. Thus, such sites are difficult to drain efficiently, and drainage sometimes significantly influences the GWL only close to the ditches, i.e. no more than about 5–15 m from them (section 3.3). A complicating factor for DNM at many peatland sites is that initial drainage usually causes subsidence of the peat (section 2.4) and, hence, increases bulk density, which aggravates drainage.
The need for DNM seems to arise earlier in northern than in southern locations, because in the north the climate is more humid and less favorable for tree growth (thus stand development is slower than in the south). Consequently, in northerly sites ditches affect stand development more strongly than in southern sites because the shift from runoff accounting for a large proportion of the water balance to evapotranspiration becoming more important is slower (cf. Sarkkola et al. 2013). This is troublesome since DNM is not always profitable in low-stocked northern stands, even if drainage conditions are poor (Ahtikoski et al. 2012).
To summarize current knowledge, four factors affect soil water conditions in a drained site: (i) the capacity of ditch networks to drain the site, (ii) the climatic conditions, mediated through the precipitation rate and potential evapotranspiration of the vegetation (tree stand and ground vegetation), (iii) the soil water conduction capacity, and (iv) the tree stand, through its water use capacity. It should be noted that DNM only affects the drainage intensity and water transportation capacity of ditch networks. However, other forest management operations (notably harvesting and fertilization) also influence stands’ water use rates. Climate cannot be influenced at all and peat’s water conductivity can be influenced only to a limited extent, in the regeneration phase by site preparation. In conclusion, as well as ditches’ condition, stand and site conditions (including the geographical location and climate), must be taken into account when assessing the need for DNM.
6.2 Applicability of the results
There have been limited numbers of studies on effects of DNM on GWL, SWC and tree growth. Therefore, this review has included some studies covering more general aspects of ditching, its effects and interactions between soil-water conditions and stand properties. Most of the results from studies in thinning-stage stands are primarily applicable to Scots pine, and to some extent Norway spruce, stands growing on drained peatlands in the Fenno-Scandinavian countries within the boreal vegetation zones (northern, middle and southern) (cf. Ahti et al. 1968). Most studies have been carried out in Finland. In Finland, and at the study sites there, the ditch systems usually consist of regularly spaced parallel ditches, while, for example in Sweden, it is common with a coulpe of main ditches within a forest stand to be drained. In the latter case, a DNM operation might be less effective, and, hence, potential growth responses smaller, when taking the whole forest stand into account. Some relevant studies conducted in North America, primarily Canada, have addressed soil-water conditions after both partial and total removal of forest stands. These studies have examined relevant phenomena for this review in both peatlands and sites with wet mineral soils, supporting other conifers than Scots pine and Norway spruce. Thus, the main findings should be quite general and applicable to many tree species and soil types within the boreal zone where forest drainage has been applied.
7 Conclusions and future research needs
7.1 Conclusions
-
Although some decade-old ditches are still functioning well, ditches gradually deteriorate over time due to subsidence of peat and blockage by vegetation, collapse of the walls, and/or accumulation of slash and other materials caused by moving machines, erosion and silting (e.g. Paavilainen and Päivänen 1995).
-
Tree stand growth may be impaired by ditch deterioration (Heikurainen 1980; Hånell 1988).
-
Ditch deterioration and subsequent impairment of tree growth are gradual processes, thus soil-water conditions may be appropriate or close to optimal for tree growth for a long time, and there is no clearly distinct optimal time for DNM.
-
There is no convenient tool for directly assessing soil-water conditions in the field, and, hence, the need for DNM in thinning-stage or older forest stands to maintain tree growth gained from initial ditching. Decisions regarding the need for DNM should not be based solely on ditch conditions, but also on stand and site properties, and geographical (climatic) factors (Sarkkola et al. 2012, 2013).
-
Visual assessment based solely on a ditch’s water transporting capacity may lead to overestimates of the need for DNM (Lauhanen et al. 1998; Laine 1986; Sarkkola et al. 2013).
-
The time since the latest ditching operation is a useful predictor of ditches’ quality. DNM will probably be needed after 20–30 years, although it may be needed any time within 10–50 years (Hökkä et al. 2000a) from ditching.
-
DNM can lower the GWL (Ahti and Päivänen 1997), thus improving soil-water conditions and, potentially, tree growth (Lauhanen and Ahti 2001; Ahti 2005; Hökkä and Kojola 2001, 2003).
-
Generally, DNM lowers the GWL by 5–10 cm in drained peatlands (Ahti and Päivänen 1997). This is similar to the GWL rise after a common operational thinning (harvest of 30–75 m3 ha–1 by stem volume; Table 1). In shallow-peated sandy subsoil sites, the GWL may be substantially lowered (Koivusalo et al. 2008).
-
On drained peatlands, available quantitative data suggest that stand growth rates will increase following DNM by 0.5–1.8 m3 ha–1 yr–1 during a 15–20-year period in Scots pine with c. 20–150 m3 ha–1 stocks. The response is higher and lasts longer in the north than in the south. For Norway spruce there is very little information available.
-
Two hypothetical requirements for increases in tree growth following DNM in drained peatlands are: (a) a pre-treatment GWL closer to the ground surface than 25–30 cm, and (b) stocking below 150 m3 ha–1. Nutrient availabilities must also be sufficient, which is not always the case in peat soils.
-
Stand stocking and management strongly affect interception, evapotranspiration, and, hence, soil-water conditions and tree growth. After timber harvesting, GWL usually rises in both peatlands and wet mineral soil sites. The reviewed studies report GWL rises of 0–15 cm after partial biomass removal and up to 40 cm after clear-cutting (Table 1).
-
The need for DMN seems to be greatest in early phases of rotations, i.e. at the time of clear-cutting and regeneration, and/or at times of pre-commercial or first commercial thinnings.
-
Drainage commonly alters peat properties by causing subsidence of the peat and increasing the degree of humification of surface layers. This implies aggravated drainage conditions in peat soils at DNM due to increased bulk density and reduced water conductivity.
-
DNM can be profitable in terms of return of investment and net present value, but not always (Aarnio et al. 1997; Hytönen and Aarnio 1998; Ahtikoski et al. 2008, 2012). Complementary ditching and a combination of complementary ditching and ditch cleaning are usually more profitable than ditch cleaning alone. Relative high initial stand stocking (c. 100–150 m3 ha–1), good–medium site quality and high temperature sum generally favor DNM profitability. Early realization of the gain in growth induced by DNM, through thinning rather than waiting until final cutting, also favors profitability.
-
Since DNM may be a harmful forestry operation from a water quality protection perspective (Joensuu et al. 2002; Nieminen et al. 2010), it is important to minimize the number of operations. A DNM operation should also result in the expected growth response, and the investment should be financially justifiable.
7.2 Research needs
Tree growth rate in poorly drained soils is not sufficiently understood, as reflected in partly conflicting and inconclusive results concerning the sufficient air filled porosity in soils for adequate tree growth. Thus, greater knowledge of effects of soil-water conditions (GWL and SWC) on tree growth are needed.
-
More empirical data on tree growth responses to DNM in drained peatlands and mineral soil sites are needed. To construct generally applicable growth models, the data should be representative for wide ranges of drained soil types, climatic conditions, and tree species (e.g. Norway spruce).
-
It is important to be able to identify when a stand is sufficiently developed to reduce excessive soil water by itself and grow at an acceptable growth rate. Inter alia further tests of the hypothesis that DNM induces weak growth responses in stands with > 150 m3 ha–1 stocks are required.
-
Further tests of the hypothesis that DNM induces weak growth responses in stands with a pre-treatment GWL of > 35–40 cm are also needed.
-
Process based models can be useful to describe and understand the interactions among all the relevant factors (ditch condition, soil properties, stand properties, and climate), quantify their impacts on soil-water conditions and tree growth in sites with peat and mineral soils, and visually display the results.
-
A convenient, robust method for assessing soil-water conditions in the field, and hence the need for, and optimal timing of, ditch maintenance to sustain tree growth at formerly drained sites.
Acknowledgements
We are grateful to Hans-Örjan Nohrstedt and Juhani Päivänen for reading a previous version of the manuscript and providing us with valuable comments and suggestions, as well as to Ari Laurén for giving some relevant advises.
References
Aarnio J., Ahti E., Hytönen L.A., Lauhanen R. (1997). Kunnostusojitus. In: Mielikäinen K., Riikilä M. (eds.). Kannattava puuntuotanto. Kustannusosakeyhtiö Metsälehti. p. 102–108. [In Finnish].
Aboal J.R., Jimnez M.S., Morales D., Gil P. (2000). Effects of thinning on throughfall in Canary Islands pine forest – the role of fog. Journal of Hydrology 238: 218–230. http://dx.doi.org/10.1016/S0022-1694(00)00329-2.
Ahti E. (1978). Maaveden energiasuhteista ojitetulla suolla. Summary: Energy relationships of soil water on drained peat. Communicationes Instituti Forestalis Fenniae 94(3): 1–56.
Ahti E. (2005). Kunnostusojitus. [Ditch network maintenance operation]. In: Ahti E., Kaunisto S., Moilanen M., Murtovaara I. (eds.). Suosta metsäksi, suometsien ekologisesti ja taloudellisesti kestävä käyttö. [From peat to forest, ecologically and economically sustainable use of peatland forests]. Tutkimusohjelman loppuraportti. Metsäntutkimuslaitoksen Tiedonantoja 947. [Finnish Forest Research Institute Research Papers 947]. p. 114–120. [In Finnish].
Ahti E., Päivänen J. (1997). Response of stand growth and water table level to maintenance of ditch networks within forest drainage areas. In: Trettin C.C., Jurgensen M.F., Grigal D.F., Gale M., Jeglum J. (eds.). Northern forested wetlands: ecology and management, chapter 32. CRC Press Inc., Lewis Publishers, USA. p. 449–457.
Ahti E., Hökkä H. (2006). Effects of the growth and volume of Scots pine stands on the level of the water table on peat in central Finland. Proceedings of an international conference on hydrology and management of forested wetlands. April 8–12 2006, New Bern, North Carolina. p. 309–315.
Ahti E., Kojola S., Nieminen M., Penttilä T., Sarkkola S. (2008). The effect of ditch cleaning and complementary ditching on the development of drained Scots pine-dominated peatland forests in Finland. In: Farrel C., Feehan J. (eds.). Proceedings of the 13th International Peat Congress. After wise use – the future of peatlands. June 8–13 2008, Tullamore, Ireland. Volume 1, Oral Presentations. International Peat Society. p. 457–459.
Ahti T., Hämet-Ahti L., Jalas J. (1968). Vegetation zones and their sections in north-western Europe. Annales Botanici Fennici 5: 169–211.
Ahtikoski A., Kojola S., Hökkä H., Penttilä T. (2008). Ditch network maintenance in peatland forest as a private investment: short- and long-term effects on financial performance at stand level. Mires and Peat 3(3). 11 p. http://mires-and-peat.net/pages/volumes/map03/map0303.php.
Ahtikoski A., Salminen H., Hökkä H., Kojola S., Penttilä T. (2012). Optimizing stand management on peatlands: the case of northern Finland. Canadian Journal of Forest Research 42: 247–259. http://dx.doi.org/10.1139/x11-174.
Bartholomeus R.P., Witte J.-P.M., van Bodegom P.M., van Dam J.C., Aerts R. (2008). Critical soil conditions for oxygen stress to plant roots: substituting the Feddes-function by a process-based model. Journal of Hydrology 360: 147– 165. http://dx.doi.org/10.1016/j.jhydrol.2008.07.029.
Berry G.J., Jeglum J.K. (1991). Hydrology of drained and undrained black spruce peatlands: groundwater table profiles and fluctuations. Canadian Forest Service, Ont. Region, Sault Ste.-Marie, Ontario. COFRDA Report 3307. 31 p. + appendice.
Boggie R., Miller H.G. (1976). Growth of Pinus contorta at different water-table levels in deep blanket peat. Forestry 49(2): 123–131. http://dx.doi.org/10.1093/forestry/49.2.123.
Brække F.H. (1983). Water table levels at different drainage intensities on deep peat in northern Norway. Forest Ecology and Management 5: 169–192. http://dx.doi.org/10.1016/0378-1127(83)90070-1.
Buss K.K., Zalitis P.P. (1968). On groundwater regime of drained forests, in Questions of Hydroforestreclamation. Zinatne Publishers, Riga. 51 p. [In Russian].
Campbell T.A. (1980). Oxygen flux measurements in organic soils. Canadian Journal of Soil Science 60: 641–650. http://dx.doi.org/10.4141/cjss80-073.
Dang Q.L., Lieffers V.J. (1989). Assessment of patterns of response of tree ring growth of black spruce following peatland drainage. Canadian Journal of Forest Research 19: 924–929. http://dx.doi.org/10.1139/a05-006.
Dubé S., Plamondon A.P. (1995). Relative importance of interception and transpiration changes causing watering-up after clear-cutting on four wet sites. Man´s influence on freshwater ecosystems and water use. Proceedings of a Boulder Symposium, July 1995. IAHS Publ. 230: 113–120.
Dubé S., Plamondon A.P., Rothwell R.L. (1995). Watering up after clear-cutting on forested wetlands of the St. Lawrence lowland. Water Resources Research 31(7): 1741–1750. http://dx.doi.org/10.1029/95WR00427.
Ernfors M., Sikström U., Nilsson M., Klemedtsson L. (2010). Effects of wood ash fertilization on forest floor greenhouse gas emissions and tree growth in nutrient poor drained peatland forests. Science of the Total Environment 408: 4580–4590. http://dx.doi.org/10.1016/j.scitotenv.2010.06.024.
Fenton N.J., Bergeron Y. (2006). Facilitative succession in a boreal bryophyte community driven by changes in available moisture and light. Journal of Vegetation Science 17(1): 65–76. http://dx.doi.org/10.1111/j.1654-1103.2006.tb02424.x.
Fraser A.I., Gardiner J.B.H. (1967). Rooting and stability in Sitka spruce. Forestry Commission Bulletin 40. HMSO, London. 28 p.
Freléchoux F., Buttler A., Schweingruber F.H., Gobat J.-M. (2000). Stand structure, invasion, and growth dynamics of bog pine (Pinus uncinata var. rotundata) in relation to peat cutting and drainage in the Jura Mountains, Switzerland. Canadian Journal of Forest Research 30(7): 1114–1126. http://dx.doi.org/10.1139/x00-039.
Glenz C., Schlaepfer R., Iorgulescu I., Kienast F. (2006). Flooding tolerance of Central European tree and shrub species. Forest Ecology and Management 235: 1–13. http://dx.doi.org/10.1016/j.foreco.2006.05.065.
Glinski J., Stepniewski W. (1985). Soil aeration and its role for plants. CRC Press, Inc., Boca Raton, FL., USA. 229 p.
Gustavsen H.G., Heinonen R., Paavilainen E., Reinikainen A. (1998). Growth and yield models for forest stands on drained peatland sites in southern Finland. Forest Ecology and Management 107: 1–17. http://dx.doi.org/10.1016/S0378-1127(97)00324-1.
Haahti K., Koivusalo H., Hökkä H., Nieminen M., Sarkkola S. (2012). Factors affecting the spatial variability of water table depth within a drained peatland forest in northern Finland. Suo 63(3–4): 107 – 121. [In Finnish with English Summary].
Hånell B. (1988). Post-drainage forest productivity of peatlands in Sweden. Canadian Journal of Forest Research 18(11): 1443–1456. http://dx.doi.org/10.1139/x88-223.
Hånell B. (1991). Shelterwood regeneration of spruce forests on productive peatlands. Swedish University of Agricultural Sciences, Department of Silviculture, Umeå. Report 32. 35 p. ISSN 0348-8969. [In Swedish with English abstract].
Heikurainen L. (1957). Metsäojien syvyyden ja pintaleveyden muuttuminen sekä ojien kunnon säilyminen. [Changes in depth and top width of forest ditches and the maintaining of their repair]. Acta Forestalia Fennica 65(3): 1–45. [In Finnish with English summary].
Heikurainen L. (1963). On using ground water table fluctuations for measuring evapotranspiration. Acta Forestalia Fennica 76(5): 1–16.
Heikurainen L. (1967). On the influence of cutting on the water economy of drained peatlands. Acta Forestalia Fennica 82(2): 1–45.
Heikurainen L. (1980). Kuivatuksen tila ja puusto 20 vuotta vanhoilla ojitusalueilla. [Drainage condition and tree stand on peatlands drained 20 years ago.] Acta Forestalia Fennica 167: 1–37. [In Finnish with English summary].
Heikurainen L., Päivänen J. (1970). The effect of thinning, clearcutting, and fertilization on the hydrology of peatland drained for forestry. Acta Forestalia Fennica 104: 1–23.
Hillman G.R. (1992). Some hydrological effects of peatland drainage in Alberta’s boreal forest. Canadian Journal of Forest Research 22: 1588–1596. http://dx.doi.org/10.1139/x92-211.
Hökkä H. (1997). Models for predicting growth and yield in drained peatland stands in Finland (Doctoral thesis). Metsäntutkimuslaitoksen tiedonantoja - The Finnish Forest Research Institute, Research Papers 651. 45 p. plus 3 appendix.
Hökkä H., Laine J. (1988). Suopuustojen rakenteen kehitys ojituksen jälkeen. [Post-drainage development of structural characteristics in peatland forest stands]. Silva Fennica 22(1): 45–65. [In Finnish with English summary].
Hökkä H., Penttilä T. (1995). Effect of thinning on groundwater table depth in drained peatlands in northern Finland. Suo 46(1): 9–19.
Hökkä H., Kojola S. (2001). Kunnostusojituksen kasvureaktioon vaikuttavat tekijät. [Factors affecting growth response due to ditch network maintenance operation]. In: Hiltunen I., Kaunisto S. (eds.). Suometsien kasvatuksen ja käytön teemapäivät. [Management and utilization of peatland forests]. Metsäntutkimuslaitoksen tiedonantoja - The Finnish Forest Research Institute, Research Papers 832. p. 30–36. [In Finnish].
Hökkä H., Kojola S. (2003). Suometsien kunnostusojitus – kasvureaktion tutkiminen ja kuvaus. [Ditch network maintenance in peatland forests – growth response and it’s description]. In: Jortikka S., Varmola M., Tapaninen S. (eds.). Soilla ja kankailla – metsien hoitoa ja kasvatusta Pohjois-Suomessa. [On peatlands and uplands – forest management in northern Finland]. Metsäntutkimuslaitoksen tiedonantoja - The Finnish Forest Research Institute, Research Papers 903. p. 13–20. [In Finnish].
Hökkä H., Alenius V., Penttilä T. (1997). Individual-tree basal area growth models for Scots pine, Norway spruce and pubescent birch on drained peatlands in Finland. Silva Fennica 31: 161–178. http://dx.doi.org/10.14214/sf.a8517.
Hökkä H., Alenius V., Salminen H. (2000a). Kunnostusojitustarpeen ennustaminen ojitusalueilla. [Predicting the need for ditch network maintenance in drained peatland sites in Finland]. Suo - Mires and Peat 51(1): 1–10.
Hökkä H., Penttilä T., Siipola M. (2000b). Spatial and temporal patterns in groundwater table level after thinning in a spruce mire. In: Rochefort L., Daigle J.-Y. (eds.). Sustaining our peatlands. Proceedings of the 11th International Peat Congress, Volume II. Canadian Society of Peat and Peatlands and International Peat Society. p. 937–942.
Hökkä H., Repola J., Laine J. (2008). Quantifying the interrelationship between tree stand growth rate and water table level in drained peatland sites within Central Finland. Canadian Journal of Forest Research 38(7): 1775–1783. http://dx.doi.org/10.1139/X08-028.
Hökkä H., Repola J., Moilanen M. (2012). Modelling volume growth response of young scots pine (Pinus sylvetris) stands to N, P, and K fertilization in drained Peatland sites in Finland. Canadian Journal of Forest Research 42: 1359–1370. http://dx.doi.org/10.1139/x2012-086.
Huikari O., Paarlahti K., Paavilainen E., Ravela H. (1966). Sarkaleveyden ja ojasyvyyden vaikutuksesta suon vesitalouteen ja valuntaan. [Summary: On the effect of stripwidth and ditch depth on the water economy and runoff on peat soil]. Communicationes Instituti Forestalis Fenniae 61(8). 39 p.
Hynynen J., Ojansuu R., Hökkä H., Siipilehto J., Salminen H., Haapala P. (2002). Models for predicting stand development in MELA System. Metsäntutkimuslaitoksen tiedonantoja - The Finnish Forest Research Institute, Research Papers 835. 116 p.
Hytönen L.A., Aarnio J. (1998). Kunnostusojituksen erilliskannattavuus muutamilla karuhkoilla rämeillä. Profitability of ditch-network maintenance on some oligotrophic pine mires. Suo 49(3): 87–99.
Isoaho P., Lauhanen R., Saarinen M. (1993). Metsäojien jatkuvan kunnossapidon vaikutus ojitusalueiden tilaan Keski-Pohjanmaalla. [Effects of continuous ditch network maintenance on the condition of forest drainage areas in Central Ostrobothnia district]. Suo 44: 33–57. [In Finnish with English abstract].
Joensuu S., Ahti E., Vuollekoski M. (2002). Effects of ditch network maintenance on the chemistry of run-off water from peatland forests. Scandinavian Journal of Forest Research 17: 238–247. http://dx.doi.org/10.1080/028275802753742909.
Jutras S., Plamondon A.P. (2005). Water table rise after harvesting in a treed fen previously drained for forestry. Suo 56(3): 95–100.
Jutras S., Plamondon A.P., Hökkä H., Bégin J. (2006). Water table changes following pre-commercial thinning on post-harvest drained wetlands. Forest Ecology and Management 235: 252–259. http://dx.doi.org/10.1016/j.foreco.2006.08.335.
Keltikangas M., Laine J., Puttonen P., Seppälä K. (1986). Vuosina 1930–1978 ojitetut suot: ojitusalueiden inventoinnin tuloksia. [Peatlands drained for forestry during 1930–1978: results from field surveys of drained areas]. Acta Forestalia Fennica 193: 1–94. [In Finnish with English summary].
Koivusalo H., Ahti E., Laurn A., Kokkonen T., Karvonen T., Nevalainen R., Finér L. (2008). Impacts of ditch cleaning on hydrological processes in a drained peatland forest. Hydrology and Earth System Sciences 12: 1211–1227. http://dx.doi.org/10.5194/hess-12-1211-2008.
Kojola S., Ahtikoski A., Hökkä H., Penttilä T. (2011). Profitability of management regimes in Scots pine stands on drained peatlands. European Journal of Forest Research. http://dx.doi.org/10.1007/s10342-011-0514-4.
Kozlowski T.T. (1982). Water supply and tree growth. Part II flooding. Review article. Forest Abstracts 43(3): 145–161.
Kukkola M., Saramäki J. (1983). Growth response in repeatedly fertilized pine and spruce stands on mineral soils. Communicationes Instituti Forestalis Fenniae 114. 55 p.
Lagergren F., Lankreijer H., Kučera J., Cienciala E., Mölder M., Lindroth A. (2008). Thinning effects on pine–spruce forest transpiration in central Sweden. Forest Ecolology and Management 255: 2312–2323. http://dx.doi.org/10.1016/j.foreco.2007.12.047.
Laine J. (1984). Estimation of evapotranspiration from peatlands by means of daily water table hydrographs. Publications from the Department of Peatland Forestry, University of Helsinki. No. 5. 100 p.
Laine J. (1986). Kuivatustekniikan, kuivatussyvyyden ja puuston kasvun välisiä vuorosuhteita 25 vuotta vanhoilla ojitusalueilla. [Interrelationships among ditching technique, ditching depth, and tree stand growth in 25 years old forest drainage areas]. Tutkimussopimushankkeen ’Metsäojitettujen soiden ekologia’ loppuraportti. Helsinki. 24 p. [In Finnish].
Landhäusser S.M., Silins U., Lieffers V.J., Liu W. (2003). Response of Populus tremuloides, Populus balsamifera, Betula papyrifera and Picea glauca seedlings to low soil temperature and water-logged soil conditions. Scandinavian Journal of Forest Research 18: 391–400. http://dx.doi.org/10.1080/02827580310015044.
Lauhanen R., Ahti E. (2001). Effects of maintaining ditch networks on the development of Scots pine stands. Suo 52(1): 29–38.
Lauhanen R., Piiroinen M.-L., Penttilä T., Kolehmainen E. (1998). Kunnostusojitustarpeen arviointi Pohjois-Suomessa. [Evaluation of the need for ditch network maintenance in northern Finland]. Suo 49(3): 101–112. [In Finnish with English summary].
Lähde E. (1969). Biological activity in some natural and drained peat soils with special reference to oxidation-reduction conditions. Acta Forestalia Fennica 94: 1–69.
Le Maitre D.C., Versfeld D.B. (1997). Forest evaporation models: relationships between stand growth and evaporation. Journal of Hydrology 193: 240–257. http://dx.doi.org/10.1016/S0022-1694(96)03144-7.
Lieffers V.J. (1988). Sphagnum and cellulose decomposition in drained and natural areas of an Alberta peatland. Canadian Journal of Soil Science 68(4): 755–761. http://dx.doi.org/10.4141/cjss88-073.
Lieffers V.J., Rothwell R.L. (1986). Effects of depth of water table and substrate temperature on root and top growth of Picea mariana and Larix lancina seedlings. Canadian Journal of Forest Research 16: 1201–1206. http://dx.doi.org/10.1139/x86-214.
Lieffers V.J., Rothwell R.L. (1987). Rooting of peatland black spruce and tamarack in relation to depth of water table. Canadian Journal of Botany 65(5): 817–821. http://dx.doi.org/10.1139/b87-111.
Lieffers V.J., MacDonald S.E. (1990). Growth and foliar nutrient status of black spruce and tamarack in relation to depth of water table in some Alberta peatlands. Canadian Journal of Forest Research 20: 805–809. http://dx.doi.org/10.1139/x90-106.
Lukkala O.J. (1949). Soiden turvekerroksen painuminen ojituksen vaikutuksesta. [Über die Setzung des Moortorfes als Folge der Entwässerungen]. Communicationes Instituti Forestalia Fenniae 37: 1–67. [In Finnish with German summary].
Lundin L. (1979). Kalhuggningens inverkan på markvattenhalt och grundvattennivån. SLU, Inst. f. Skoglig Marklära. Rapporter i skogsekologi och skoglig marklära. 35 p. ISBN 9157603308. [In Swedish with English summary].
Lundin L. (1999). Effects on hydrology and surface water chemistry of regeneration cuttings in peatland forests. International Peat Journal 9: 118–126.
Lundin L. (2000). Water environment care at peatland forestry practices. In: Rochefort L., Daigle J.-Y. (eds.). Sustaining our peatlands. Proceedings of the 11th International Peat Congress, Volume II. Canadian Society of Peat and Peatlands and International Peat Society. p. 952–961.
Mannerkoski H., Finér L., Piirainen S., Starr M. (2005). Effect of clear-cutting and site preparation on the level and quality of groundwater in some headwater catchments in eastern Finland. Forest Ecology and Management 220: 107–117. http://dx.doi.org/10.1016/j.foreco.2005.08.008.
Marcotte P. (2005). Rétablissement hydrologique après coupe et vigeur de la regeneration naturelle en sapin baumier (Abies balsamea (L.) Mill.) de milieu forestiers humides de la vallée du St. Laurent. In: Mémoire de mâitrise. Faculté de foresterie et de géomatique, Université Laval, Sainte-Foy, Quebeq. 60 p.
Marcotte P., Roy V., Plamondon A.P., Auger I. (2008). Ten-year water table recovery after clearcutting and draining boreal forested wetlands of eastern Canada. Hydrological Processes 22: 4163–4172. http://dx.doi.org/10.1002/hyp.7020.
McDonald S.E., Yin F. (1999). Factors influencing size inequality in peatland black spruce and tamarack: evidence from post-drainage release growth. Journal of Ecology 87: 404–412. http://dx.doi.org/10.1046/j.1365–2745.1999.00370.x.
Minkkinen K., Laine J. (1998a). Long-term effects of forest drainage on the peat carbon stores of pine mires in Finland. Canadian Journal of Forest Research 28: 1267–1275. http://dx.doi.org/10.1139/x98-104.
Minkkinen K., Laine J. (1998b). Effects of forest drainage on the peat bulk density of pine mires in Finland. Canadian Journal of Forest Research 28: 178–186. http://dx.doi.org/10.1139/x97-206.
Minkkinen K., Byrne K.A., Trettin C. (2008). In: Strack M. (ed.). Peatland and climate change, chapter 4. International Peat Society. p. 98–122. ISBN 978-952-99401-1-0.
Mäkitalo K., Hyvönen J. (2004). Late-summer soil water content on clear-cut reforestation areas two decades after site preparation in Finnish Lapland. Forest Ecology and Management 189: 57–75. http://dx.doi.org/10.1016/j.foreco.2003.07.036.
Mäkitalo K., Heiskanen J., Hallikainen V. (2006). Long-term effect of ploughing on soil hydrology in northern Finland. In: Proceedings of an international conference on hydrology and management of forested wetlands, April 8–12 2006, New Bern, North Carolina, p. 284–291.
National Forest Program 2015. Forest sector as the responsible actor of bioeconomy (2011). Maa- ja metsätalousministeriö. [Kansallinen metsäohjelma 2015. Metsäalasta biotalouden vastuullinen edelläkävijä]. ISBN 978-952-453-630-1. [In Finnish].
Nieminen M., Ahti E., Koivusalo H., Mattsson T., Sarkkola S., Laurén A. (2010). Export of suspended solids and dissolved elements from peatland areas after ditch network maintenance in south-central Finland. Silva Fennica 44(1): 39–49. http://dx.doi.org/10.14214/sf.161.
Olkinuora M. (1990). Kunnostusojituksen vaikutus pohjavesipinnan tasoon ja puuston kasvuun. MSc Thesis, Univ. Helsinki, Dep. Peatland For., Helsinki, Finland. 54 p.
Paavilainen E. (1967). Männyn juuriston suhteesta turpeen ilmatilaan. Summary: Relationships between the root system of Scots pine and the air content of peat. Communicationes Instituti Forestalis Fenniae 63(3): 1–21.
Paavilainen E., Päivänen J. (1995). Peatland forestry: ecology and priciples. Ecological Studies 111. 248 p. Springer-Verlag, Berlin-Heidelberg-New York. ISBN 3-540-58252-5.
Payandeh B. (1973). Analyses of forest drainage experiment in northern Ontario, I: Growth analysis. Canadian Journal of Forest Research 3: 387–398.
Päivänen J. (1966). Sateen jakaantuminen erilaisissa metsiköissä. [The distribution of rainfall in different types of forest stands]. Silva Fennica 119(3): 1–37. [In Finnish with English summary]. http://dx.doi.org/10.14214/sf.a14290.
Päivänen J. (1973). Hydraulic conductivity and water retention in peat soils. Acta Forestalia Fennica 129. 70 p.
Päivänen J. (1980). The effect of silvicultural treatments on the hydrology of old forest drainage areas on peat. Proceedings of the Helsinki symposium “The influence of man on the hydrological regime with special reference to representative and experimental basins”, 23–26 June 1980. IAHS-AISH Publication 130: 137–140.
Päivänen J. (1982). Hakkuun ja lannoituksen vaikutus vanhan metsäojitusalueen vesitalouteen. [Summary: The effect of cutting and fertilization on the hydrology of an old forest drainage area]. Folia Forestalia 516: 1–19.
Päivänen J. (1984). The effect of runoff regulation on tree growth on a forest drainage area. Proceedings 7th International Peat Congress, vol. 3: 476–488. Irish Nat Peat Comm, Dublin.
Päivänen J. (1990). Suometsät ja niiden hoito. [Peatland forestes and their management]. Kirjayhtymä, Helsinki. 231 p. [In Finnish].
Päivänen J., Ahti E. (1988). Ditch cleaning and additional ditching in peatland forestry – effect on ground water level. Proceedings of the international symposium on the hydrology of wetlands in temperate and cold regions. Joensuu, Finland, 6–8 June 1988. Publications of the Academy of Finland 4: 184–189.
Päivanen J., Sarkkola S. (2000). The effect of thinning and ditch network maintenance on the water table level in a Scots pine stand on peat soil. Suo 51(3): 131–138.
Päivänen J., Hånell B. (2012). Peatland ecology and forestry – a sound approach. University of Helsinki Department of Forest Sciences Publications 3: 1–267. ISBN 978-952-10-4531-8.
Pelkonen E. (1975). Vuoden eri aikoina korkella olevan pohjaveden vaikutus männyn kasvuun. [Effects on Scots pine growth of ground water adjusted to the ground surface for periods of varying length during different seasons of the year]. Suo 26(2): 25–32.
Pettersson F. (1994). Predictive functions for impact of nitrogen fertilization on growth over five years. The Forestry Research Institute of Sweden, Report No. 3. Uppsala. 56 p. ISSN 1103–6648.
Plamondon A.P., Prévost M., Naud R.C. (1984). Interception de la pluie dans la sapinièr à bouleau blanc, Forêt Montmorency. Canadian Journal of Forest Research 14: 722–730. Abstract. http://dx.doi.org/10.1139/x84-129.
Ponnamperuma F.N. (1984). Effects of flooding on soil. Chapter II. In: Kozlowski T.T. (ed.). Flooding and plant growth. Academic Press. Inc. London Ltd. p. 9–42. ISBN 0-12-424120-4.
Porthier D., Prévost M., Auger I. (2003). Using the shelterwood method to mitigate water table rise after forest harvest. Forest Ecology and Management 179: 573–583. http://dx.doi.org/10.1016/S0378-1127(02)00530-3.
Prévost M., Gauthier M.-M. (2013). Shelterwood cutting in red spruce – balsam fir lowland site: effects of final cut on water table and regeneration development. Forest Ecology and Management 291: 404–416. http://dx.doi.org/10.1016/j.foreco.2012.11.019.
Price J.S., Heathwaite A.L., Baird A.J. (2003). Hydrological processes in abandoned and restored peatlands: an overview of management approaches. Wetlands Ecology and Management 11(1–2): 65–83. http://dx.doi.org/10.1023/A:1022046409485.
Rosen M. (1989). Forest drainage manual. Ontario Ministry of Natural Resources. Science and Technology Series 3: 1–25.
Rothwell R.L., Silins U., Hillman. G.R. (1996). The effect of drainage on substrate water content at several forested Alberta peatlands. Canadian Journal of Forest Research 26: 53–62. http://dx.doi.org/10.1139/x26-006.
Roy V., Jeglum J.K., André P.P. (1997). Water table fluctuations following clearcutting and thinning on Wally creek wetlands. In: Tretton C.C., Jurgensen M.F., Grigal D.F., Gale M., Jeglum J. (eds.). Northern forested wetlands: ecology and management. CRS Press Inc., Lewis Publishers, USA. p. 239–251.
Roy V., Plamondon A.P., Bernier P.-Y. (2000a). Draining forested wetland cutovers to improve seedling root zone conditions. Scandinavian Journal of Forest Research 15: 58–67. http://dx.doi.org/10.1080/02827580050160475.
Roy V., Ruel J.-C., Plamondon A.P. (2000b). Establishment, growth and survival of natural regeneration after clearcutting and drainage on forested wetlands. Forest Ecolology and Management 129: 253–267. http://dx.doi.org/10.1016/S0378-1127(99)00170-X.
Saarinen M., Silver T., Joensuu S. (1998). Ojien mitoitus kunnostusojituksessa. Kirjallisuustarkastelu. [Ditch dimensioning in ditch-network-maintenance areas – a literature review]. Suo 49(3): 75–85. [In Finnish with English summary].
Sarkkola S., Hökkä H., Koivusalo H., Nieminen M., Ahti E., Päivänen J., Laine J. (2010). Role of tree stand evapotranspiration in maintaining satisfactory drainage conditions in drained peatlands. Canadian Journal of Forest Research 40: 1485–1496. http://dx.doi.org/10.1139/X10-084.
Sarkkola S., Hökkä H., Ahti E., Koivusalo H., Nieminen M. (2012). Depth of water table prior to ditch network maintenance is a key factor for tree growth response. Scandinavian Journal of Forest Research 27: 649–658. http://dx.doi.org/10.1080/02827581.2012.689004.
Sarkkola S., Nieminen M., Koivusalo H., Laurén A., Ahti E., Launiainen S., Nikinmaan E., Marttila H., Laine J., Hökkä H. (2013). Domination of growing-season evapotranspiration over runoff makes ditch network maintenance in mature peatland forests questionable. Mires and Peat 11(2): 1–11. http://www.mires-and-peat.net/pages/volumes/map11/map1102.php.
Seppälä K. (1969). Kuusen ja männyn kasvun kehitys ojitetuilla turvemailla. [Post-drainage growth rate of Norway spruce and Scots pine on peat]. Acta Forestalia Fennica 93: 5–88. [In Finnish with English summary].
Silins U. (1997). Post-drainage peatland moisture and aeration dynamics. Ph.D. thesis. University of Alberta, Edmonton.
Simard M., Lecomte N., Bergeron Y., Bernier P.Y., Pare D. (2007). Forest productivity decline caused by successional paludification of boreal soils. Ecological Applications 17: 1619–1637. http://dx.doi.org/10.1890/06-1795.1.
Stewart G.G. (1958). Preliminary results of experiments in drain deepening in two Border forests. Reproduced from Rep. For. Res. for the year ending of March 1958. p 131–137.
Sutton R.F. (1993). Mounding site preparation: a review of European and North American experience. New forests 7: 151–192. http://dx.doi.org/10.1007/BF00034198.
Timonen E. (1983). Havaintoja auraus- ja kaivuriojien mitoista ja kunnosta soilla. [The size and the condition of ditches made by ploughs and tractor diggers in drained peatlands]. Suo 34: 29–39. [In Finnish with English summary].
Toth J., Gillard D. (1988). Experimental design and evaluation of a peatland drainage system for forestry by optimization of synthetic hydrographs. Canadian Journal of Forest Research 18: 353–373. http://dx.doi.org/10.1139/x88-053.
Trettin C. (1986). Application of water management techniques to hardwood silviculture. In: Mroz G., Reed D. (eds.). Proceedings of a conference on the northern hardwood resource: management and potential, August 18–30, 1986. Michigan Technological University, Houghton, Michigan. p. 183–192.
Vincke C., Thiry Y. (2008). Water table is a relevant source for water uptake by a Scots pine (Pinus sylvestris L.) stand: evidence from continuous evapotranspiration and water table monitoring. Agricultural and Forest Meteorology 148: 1419–1432. http://dx.doi.org/10.1016/j.agrformet.2008.04.009.
Vompersky S.E., Sabo E.D., Formin A.S. (1975). Forest drainage amelioration. Lesnaja Promyshlennost Publishers, Moscow. 295 p. [In Russian].
Vompersky S., Sirin A. (1997). Hydrology of drained forested wetlands. In: Trettin C.C., Jurgensen M.F., Grigal D.F., Gale M.R., Jeglum J.K. (eds.). Nothern forested wetlands. Ecology and management. CRC Press, Lewis Publishers. p. 189–211.
Wang A.-F., Roitto M., Lehto T., Zwiazek J.J., Calvo-Polanco M., Repo T. (2013). Waterlogging under simulated late-winter conditions had little impact on the physiology and growth of Norway spruce seedlings. Annals of Forest Science 70: 781–790. http://dx.doi.org/10.1007/s13595-013-0325-5.
Wesseling J., van Wijk W.R. (1957). Soil physical conditions in relation to drain depth. In: Luthin J.N. (ed.). Drainage of Agricultural Lands. American Society of Agronomy, Madison, Wisconson. p. 461–504.
Total of 126 references.
