Lars Karlsson  ,
Tommy Mörling,
Urban Bergsten
,
Tommy Mörling,
Urban Bergsten
Influence of silvicultural regimes on the volume and proportion of juvenile and mature wood in boreal Scots pine
Karlsson L., Mörling T., Bergsten U. (2013). Influence of silvicultural regimes on the volume and proportion of juvenile and mature wood in boreal Scots pine. Silva Fennica vol. 47 no. 4 article id 938. https://doi.org/10.14214/sf.938
Highlights
- Initial stand densities have a large impact on the proportion of mature wood within trees throughout most of their life cycle
- The difference between regimes in volume of long fibres in crop trees could be substantial
- Long-term silvicultural strategies implemented at juvenile stand ages can be important tools in order to produce wood raw material suited for specific end-uses.
Abstract
Trees from 48 to 78 years old, exposed to three different long-term silvicultural regimes, were examined for transition ages between juvenile (JW), transition (TW) and mature wood (MW), total wood volume and proportions of the same wood types, as defined by fibre length. Twenty one sample trees were collected at sites with similar growing conditions within the same geographical area. Stem discs and fibre samples from breast height (BRH), 20% of tree height, green crown height and 70% of tree height were analysed. Trees growing in an environment with few neighboring trees (Sparse regime) started to produce MW, on average, five years later at BRH and six to nine years later at 20% of total tree height than trees in stands with high stem numbers (Dense regime) and trees growing in stands where the stem number had been heavily reduced after an initial high stand density (Dense/Sparse regime). For all regimes, the greatest mean fibre length was found below the green crown and high initial stem numbers were found to positively influence fibre length. The proportion of MW in the whole stem was 34% at Sparse regime sites, 57–69% at Dense/Sparse sites and 63–64% in sites where there was a Dense regime. The proportion of MW was significantly lower for trees from the Sparse regime on each stem section compared. In conclusion, the results suggest that the initial condition a tree faces affects the stem wood properties and quality output at the end of the rotation period.
Keywords
fibre length;
stand density;
juvenile wood
-
Karlsson,
Department of Forest Biomaterials and Technology, Swedish University of Agricultural Sciences, SE-901 83 Umeå, Sweden
E-mail
lars.karlsson@slu.se
- Mörling, Department of Forest Ecology and Management, Swedish University of Agricultural Sciences, SE-901 83 Umeå, Sweden E-mail tommy.morling@slu.se
- Bergsten, Department of Forest Biomaterials and Technology, Swedish University of Agricultural Sciences, SE-901 83 Umeå, Sweden E-mail urban.bergsten@slu.se
Received 14 June 2013 Accepted 12 November 2013 Published 3 December 2013
Views 168354
Available at https://doi.org/10.14214/sf.938 | Download PDF

1 Introduction
Commercial forestry in Fennoscandia generally involves regeneration measures, pre-commercial thinning (PCT) to sparse stand stem densities, one or several thinnings and finally a clear-cutting. Although this regime is prevalent in Swedish forestry, the area of young, dense forests is increasing rather rapidly (Swedish Statistical Yearbook 2011), calling for alternative views on forest management regimes. Bioenergy could be derived from dense young forests in a first fuelwood thinning. It is important when harvesting dense stands to retain a higher basal area and stem number than in a conventional first thinning, in order to avoid production losses (Nilsson et al. 2010). In this way, stands are kept dense for a substantially longer time than in conventional management regimes.
One major concern associated with thinning in young dense stands has been that little is known about the further wood properties and quality development of the remaining trees. In general, wood characteristics are the result of the interaction between tree age, genetic make-up and environmental influence. Although quality traits associated with stand density have been frequently reported, most studies seem to focus on the effects of single treatments rather than those of long-term regimes, using several silvicultural measures over time. For instance, when compared with widely-spaced stands, dense stands generally produce trees with a small microfibril angle (Eriksson et al. 2006), a high relative density (Yang and Hazenberg 1994; Yang 2002), a low occurrence of knots and tapering (Persson 1977) and small branch diameters at the lower part of the stem (Johansson 1992; Varmola and Salminen 2004; Fahlvik et al. 2005).
Sauter et al. (1999) and Ulvcrona and Ulvcrona (2011) have suggested that studies of wood should also include potential effects on juvenile wood (JW). JW refers to the innermost section of wood within a tree stem. The formation of JW has been explained as being caused by several processes, often involving auxin production (Zobel and Sprague 1998). As wood properties gradually change with age, there is no definite point where the JW zone ends and the mature wood (MW) zone begins. The transition age (i.e. the transition zone) has been defined as the change in magnitude of one or several anatomical features in relation to the distance to the pith (Dinwoodie 2000). Thus, three different wood types can be defined, namely JW, transition wood (TW) and MW (Boutelje 1968; Briggs and Smith 1986). The determination of JW will thus depend on the definition and anatomic feature addressed. In order to define and determine JW proportion, features such as density (Clark and Saucier 1989; Kucera 1994; Sauter et al. 1999; Mutz et al. 2004; Gapare et al. 2006), fibre length (Yang et al. 1986; Kucera 1994; Yang 1994; Yang and Hazenberg 1994; Fries et al. 2003), ring area (Alteyrac et al. 2006) and longitudinal shrinkage (Saranpää 1994) have all previously been used. However, fibre length could be the most consistent parameter defining transition age between JW and MW since fibre traits could be directly linked to growth trends according to Mansfield et al. (2009).
The age interval at which JW is formed often varies between individual trees and between tree species (Panshin and de Zeeuw 1980). The transition age between JW and MW has been investigated and reported for several tree species, including: Larix kaempferi 18 years (Zhu et al. 2000), Picea abies 5–29 years (Kucera 1994; Saranpää 1994), Picea glauca 12–16 years, Picea mariana 12–21 years (Yang 1994; Yang and Hazenberg 1994; Alteyrac et al. 2006), Pinus contorta 13–25 years (Mansfield et al. 2009), Pinus eliottii 10 years (Clark and Saucier 1989), Pinus radiata 5–22 years (Gapare et al. 2006), Pinus sylvestris 16–27 years (Sauter et al. 1999; Mutz et al. 2004; Fries et al. 2003) and Pinus taeda 12–14 years (Bendtsen and Senft 1986; Clark and Saucier 1989). Therefore, a general guideline often used is that JW in conifers includes the first 5–25 growth rings. Typically, JW is more apparent in conifers than in deciduous tree species (Bendtsen and Senft 1986) with higher proportions found in plantation forests compared to natural regenerated stands (Bendtsen 1978). Moreover, the length of the growing season (Clark and Saucier 1989) and genetic control (Abdel-Gadir and Krahmer 1993; Gapare et al. 2006) have been found to influence the demarcation between JW and MW.
In general, information about the characteristics of raw material facilitates the process of optimizing the utilization of the output from active forest management and thus increases the value of wood raw material. For JW, a short tracheid length (Panshin and de Zeeuw 1980; Harris 1981; Bendtsen and Senft 1986; Yang and Hazenberg 1994), a large microfibril angle (Harris 1981; Bendtsen and Senft 1986) and low wood density (Pearson and Gilmour 1971; Harris 1981; Bendtsen and Senft 1986) act together and negatively affect the mechanical properties of coniferous trees. Due to the large fibril angle, JW tends to shrink in the longitudinal direction after drying (Pearson and Gilmour 1971; Saranpää 1994). Therefore, solid wood products containing JW might be unstable. Moreover, a tree’s compression wood content is normally heavily concentrated in the JW zone (Bendtsen and Senft 1986). The high lignin content, the low amount of cellulose and the high moisture content in JW (Uprichard and Lloyd 1980) lead to a complicated and expensive pulping process. The chemical content of JW is mainly due to fibre morphology and the thickness of the cell wall (Uprichard and Lloyd 1980). Chemical pulp yields per unit volume have been reported to be reduced by 5–15% when JW is used (Kirk et al. 1972; Zobel and Sprague 1998). The short JW fibres with thin cell walls produce pulp with low tear strength properties, low opacity, high tensile and burst strength (Kirk et al. 1972). In the thermo-mechanical pulping process (TMP), JW consumes more electricity than MW but produces paper with good strength and optical properties (Myers 2001). Products such as OSB (oriented strand board), MDF (medium density fibreboard), TMP and CTMP (chemithermomechanical pulp) can all be made from JW (Zobel and Sprague 1998).
The current interest in deriving biomass from dense forest stands means that stem wood quality must be monitored so that the subsequent production of pulp and timber is not jeopardized. In the process of characterizing the overall quality output of a management regime, the volume and the proportion of JW serve as useful indicators. This study therefore aims to clarify whether the transitions between juvenile wood, transition wood and mature wood are affected by different silvicultural practices in different rotation phases. We hypothesize that the initial environment trees face has long-lasting effects on the quality of the wood produced. Thus, we have tested to see if the initial conditions (e.g. stand density) affect the length of time before fibres of mature length are formed within a tree and what long-term effects could be expected. Therefore, the objective of this study was to quantify and compare wood volumes of different fibre length classes within the stem of trees in (i) single different treatments and (ii) different conceptual management regimes (i.e. a long-term silvicultural strategy) that might be considered in practical forestry.
2 Material and methods
2.1 Study areas, treatments and regimes
2.1.1 Study areas
The Swedish University of Agricultural Science´s database of long-term silvicultural trials (Karlsson and Ulvcrona 2010) was searched for stands that had been subjected to three different management regime types. Suitable stand types with a similar site index (SI; Hägglund 1974) and comparable altitude levels within the same geographical region were chosen (Table 1). Treatments of various magnitudes were chosen to be included in the various management regimes.
| Table 1. Geographical position, altitude (meters above sea level), site index (Hägglund 1974) and year of stand establishment of the study sites used to compare silvicultural regime types (Dense/Sparse = a high initial stand density followed by high intensity pre-commercial thinning/commercial thinning, Dense = a continuous high stand density and Sparse = a continuous low stand density). | |||||
| Regime type | Site name | Geographical position | Alt. (m.a.s.l.) | SI (H100) | Year of stand establishment |
| Dense/Sparse | Svartberget (SVA) | 64°14´N, 19°46´E | 210 | 23 | 1948 |
| Svartberget | 64°14´N, 19°46´E | 210 | 23 | 1948 | |
| Norrliden (NOR) | 64°21´N, 19°46´E | 240 | 21 | 1956 | |
| Dense | Svartberget | 64°14´N, 19°46´E | 210 | 23 | 1948 |
| Kulbäcksliden (KUL) | 64°10´N, 19°35´E | 260 | 22 | 1934 | |
| Sparse | Åsele (ÅSE) | 64°14´N, 17°33´E | 330 | 23 | 1964 |
| Åheden (ÅHE) | 64°09´N, 19°40´E | 195 | 19 | 1948 | |
2.1.2 Dense regime
Sample trees representing a management regime where the trees had been growing within a densely populated stand throughout their life were collected from two different treatments at the sites at Kulbäcksliden and Svartberget (Table 1).
The treatments were:
- Natural regeneration, no PCT and no thinning until age 46 (thinning to 1400 stems ha–1) when the dominant height was 12.5 m (No PCT; NP)
- Natural regeneration + planting (1.7 m spacing), no PCT until age 23 (when PCT to 1500 stems ha–1 was carried out) and no thinning during the following observed 42 years (No thinning; NT).
2.1.3 Dense/Sparse regime
Three different types of treatment at the sites at Svartberget and Norrliden (Table 1) were chosen as representative of trees exposed to a regime where trees initially experienced high competition from neighboring trees and later, after PCT and/or thinning, grew without trees in their immediate vicinity.
The treatments were:
- Natural regeneration + planting (1.7 m spacing), PCT to 1500 stems ha–1 at age 23 and thinning at age 50 leaving (i) the 300 largest stems ha–1 (300L) and (ii) the 300 smallest stems ha–1 (300S) for free development during 16 years.
- Natural regeneration, PCT to 600 stems ha–1 at age 24 when average stand height was 6 m, no thinning (P600) over the following 32 years.
2.1.4 Sparse regime
Sample trees were collected from two different treatments at locations Åsele and Åheden (Table 1).
The treatments were:
- Planting and maintenance of 3 m spacing (3M) over a period of 44 years
- Planting (3 m spacing) carried out under a very sparse stand density of seed/shelter trees. Harvesting of seed/shelter trees 5 years after planting and PCT with herbicide treatments 5–9 years after establishment leaving trees growing in a spacing of 10 m (10M) over a period of 55 years.
2.2 Sampling
Three sample trees were chosen randomly within the dominant- and co-dominant tree classes of each treatment. Detailed information about all 21 sample trees and their characteristics are presented in Table 2. In order to model whole-tree proportions and volumes of JW, TW and MW stem discs were removed along the stem. For every sample tree, 20–30 cm thick stem discs were removed at breast height (1.3 m), at the lowest height of the living crown and at 20% and 70% of the total tree height. Stem discs were measured for diameter, number of rings, width of individual growth rings and proportions of earlywood and latewood, using a commercially available scanner and the software package WinDENDROTM. From the stem discs, two approximately 3 mm thick wood sticks were sawn in the radial direction (from pith to bark) in the north-south direction of the trees. The sticks were divided in the longitudinal direction at every third year ring, counting from the pith and moving outwards. As the main objective was to find the age at which MW was first produced, it was not considered necessary to prepare specimens outside ring number 36. Thus, specimens containing three annual rings were taken in order to represent different cambial ages within the first 36 year rings.
| Table 2. Characteristics of all sample trees included in the study. Site abbreviations are explained in Table 1. DBH = diameter at breast height; DGC = diameter at the lowest living branch (not surrounded by two dead branch whorls) of the green crown; D70 = diameter at 70% of total tree height. Dense/Sparse = a high initial stand density followed by high intensity pre-commercial thinning/commercial thinning, Dense = a continuous high stand density, and Sparse = a continuous low stand density. 300L = thinning leaving the 300 largest trees ha–1; 300S = thinning leaving the 300 smallest trees ha–1; P600 = PCT to 600 stems ha–1 at a dominant height of 5m; NT = no thinning; NP = no PCT; 3M = 3 m spacing; 10M = 10 m spacing. | |||||||||
| Regime type | Treatment | Site | No. of year rings at breast height | Height (m) | Height to living crown (m) | Height to living crown (% of tree height) | DBH (cm) | DGC (cm) | D70 (cm) |
| Dense/ Sparse | 300S | SVA | 46 | 14.3 | 6.1 | 42.7 | 19.3 | 14.6 | 9.7 |
| SVA | 47 | 14.7 | 7.6 | 51.7 | 18.2 | 13.0 | 9.1 | ||
| SVA | 46 | 12.1 | 6.7 | 55.4 | 15.0 | 10.2 | 8.1 | ||
| 300L | SVA | 50 | 16.7 | 5.5 | 32.9 | 23.8 | 20.4 | 11.1 | |
| SVA | 52 | 17.1 | 6.4 | 37.4 | 27.0 | 20.5 | 12.9 | ||
| SVA | 46 | 14.1 | 6.0 | 42.5 | 20.5 | 16.6 | 10.3 | ||
| P600 | NOR | 42 | 17.6 | 7.2 | 40.9 | 22.4 | 18.5 | 11.6 | |
| NOR | 43 | 18.6 | 9.9 | 53.2 | 18.5 | 12.5 | 9.1 | ||
| NOR | 43 | 17.6 | 8.9 | 50.6 | 20.3 | 14.5 | 10.2 | ||
| Dense | NT | SVA | 48 | 18.6 | 10.5 | 56.4 | 16.7 | 9.7 | 7.7 |
| SVA | 47 | 15.9 | 8.5 | 53.5 | 14.1 | 9.5 | 6.9 | ||
| SVA | 46 | 17.0 | 9.0 | 52.9 | 16.2 | 10.5 | 8.1 | ||
| NP | KUL | 69 | 20.7 | 13.9 | 67.1 | 19.6 | 13.2 | 11.4 | |
| KUL | 64 | 18.2 | 12.5 | 68.7 | 15.2 | 8.0 | 7.9 | ||
| KUL | 65 | 19.2 | 12.3 | 64.1 | 18.5 | 9.6 | 8.5 | ||
| Sparse | 3 M | ÅSE | 36 | 13.9 | 5.7 | 41.0 | 17.5 | 14.1 | 8.6 |
| ÅSE | 37 | 15.2 | 5.8 | 38.2 | 22.2 | 16.5 | 9.8 | ||
| ÅSE | 38 | 14.5 | 6.7 | 46.2 | 18.1 | 13.1 | 9.9 | ||
| 10M | ÅHE | 43 | 15.2 | 3.5 | 23.0 | 29.3 | 26.6 | 10.8 | |
| ÅHE | 43 | 12.9 | 4.5 | 34.9 | 22.0 | 14.2 | 7.7 | ||
| ÅHE | 45 | 13.9 | 4.2 | 30.2 | 27.0 | 20.5 | 10.0 | ||
2.3 Fibre analysis
All specimens were visually inspected for occurrence of compression wood. In the cases of obvious occurrence, specimens were not further analyzed. Fibre extraction and removal of wood components were carried out in accordance with the work by Franklin (1945) and Fries et al. (2003). Specimens were placed in test tubes with a mixture of equal volumes of hydrogen peroxide (H2O2) diluted to 25% and concentrated acetic acid (CH3COOH). Test tubes were stored in an oven (70 °C) until the wood was pale, which took approximately 24–30 hours. Once the specimens were removed from the oven, they were washed three times in water before being shaken until a homogenous fibre suspension was formed. The suspensions were analyzed using a Kajaani FiberLab 3.0 analyzer (Metso Automation Inc., Kajaani, Finland). Between every run, each piece of equipment was rinsed in order to avoid sample contamination. For calibration, every tenth specimen was analyzed three times. In order to eliminate the influence of small fragments, length-square-weighted mean length (Fries et al. 2003) was calculated as:
![]()
where n is the number of fibres in each length class and li is the mid-class length in class i. After visual examination of pictures taken during measuring, the maximum fibre length was set to 4.0 mm and the minimum length was set to 0.3 mm.
2.4 Calculations
2.4.1 Determination of transition age between fibre length classes
Functions expressing fibre length development over time were fitted in the interval of 0.3–2.8 mm. For simplicity, in order to keep the number of function expressions to a workable level, two different expressions were used:
![]()
![]()
where y is the fibre length, z is the growth ring number (counted from the pith and outwards), a is the intercept and b and c represent the slope of the expression. Suitable expression was determined using the standard deviation of the fitted line.
The measurements of each tree were divided into three fibre length classes: < 1.5 mm, 1.5–2.5 mm and > 2.5 mm representing JW, TW and MW respectively (Boutelje 1968). Transition ages at 1.5 mm and 2.5 mm were found by setting y equal to 1.5 and 2.5 in the expression used for fibre length development (2 or 3). Then x was derived as:
![]()

For each individual tree, the arithmetic means of fibre length were calculated within each of the three defined length classes. Thus, it was assumed that the fibre length would not increase (or decrease) substantially outside the sample area. Total mean fibre length (![]() ) was then calculated as:
) was then calculated as:
![]()
where ![]() is the arithmetic mean fibre length within fiber length class i for tree j at height h and P is the proportion of total tree radius.
is the arithmetic mean fibre length within fiber length class i for tree j at height h and P is the proportion of total tree radius.
2.4.2 Volumes and volume proportions of fibre length classes
In order to calculate the volume and volume proportion of each fibre length class, the trees were divided into five sections. The sampling heights constituted the boundary between different sections. Thus, the stem sections were: 0–1.3 m (Base), 1.3 m to 20% of total tree height (Stem 1), 20% of total tree height to the height of the living crown (Stem 2), height of living crown to 70% of total tree height (Stem 3), and 70% to 100% of total tree height (Top). Calculations were largely in accordance with the work of Alteyrac et al. (2006), although some modifications were made. The radii of fibre length classes (FLCR) were calculated as:

where TAijh is the transition age at fibre length i in tree j at sampling height h and RW is the ring width. Two different transition ages were used: i = 1.5 mm and i = 2.5 mm.
Tree radius (TR) was calculated as:

where RW is the ring width at cambial age a. The formula for a truncated cone was used to calculate fibre length class volume (FLC_Vol_Stemijh) for class i and total volumes (Tot_Vol_Stemjh) for sections Stem 1, Stem 2, and Stem 3. The volume of stemijh is the part of the stem from the h–1th sampling height to the hth sampling height.
![]()
![]()
The formula for a cone was used to calculate fibre length class volume (FLC_Vol_Topijh) and total volume (Tot_Vol_Topjh) in the Top stem section.
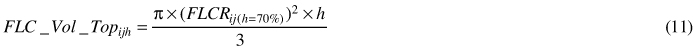

The formula for a cylinder was used to calculate fibre length class volume (FLC_Vol_Baseijh) and total volume (Tot_Vol_Basejh) in the Base stem section (0–1.3 m).
![]()
![]()
Total tree volume (TotVol) and total volume in fibre length class i (FLCVol) were calculated as:
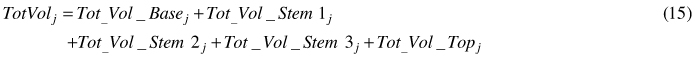
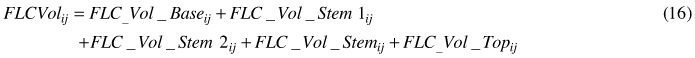
For i = 1.5–2.5 mm (TW zone), the volume was calculated as:
![]()
For i = > 2.5 mm (MW zone), the volume was calculated as:
![]()
Finally, the volume proportion of fibre length class i (FLCpropVolij) was calculated as:

2.5 Statistical analysis
Mean fibre length and ages of demarcation between the identified fibre length classes were analyzed at the different sample heights; wood type volume ha–1 and wood type volume proportions were also compared. Treatments within the same regime were compared in order to reveal if they differed significantly and if the division into regimes was relevant. Thereafter, comparisons were made, first between treatments and, second, between regimes, using the GLM procedure in the MINITAB statistical software (Minitab Statistical Software 2007). Significant differences (p ≤ 0.05) were analyzed using Tukey’s test.
3 Results
3.1 Transition ages
The analysis of variance revealed statistically significant differences in transition age below the green crown between regime type and also between treatments (Table 3). Most treatments started to produce 1.5 mm long fibres at about the same age. However, the results at breast height and at 20% of total tree height indicate that the 1.5 mm length was reached at a relatively late age for treatments which comprised planting at a sparse spacing. However, the only statistically significant difference in this respect was found at breast height between treatments 10M and P600. At breast height and at 20% of total tree height, 1.5 mm long fibres were produced within 4.0 (300S) to 10.8 (10M) years. The mature fibre length (2.5 mm) was reached within 17.1 (P600) to 28.2 (10M) years at breast height and within 12.9 (NT) to 22.5 (10M) years at 20% of total tree height (Table 4). For all treatments, the mature state was reached at a lower age at 20% of total tree height compared to breast height. Significant statistical differences between treatments at 20% were detected between 10M which differed from both NT and NP in terms of producing fibres of mature length (Table 4).
| Table 3. Effects of treatments and regimes on transition ages between different fibre length classes at different tree heights, as shown by analysis of variance. | ||||||
| Variable | Treatment | Regime | ||||
| df | F | P | df | F | P | |
| Breast height | ||||||
| Age at 1.5 mm | 6 | 3.15 | 0.036 | 2 | 5.13 | 0.017 |
| Age at 2.5 mm | 6 | 1.81 | 0.169 | 2 | 3.27 | 0.062 |
| Mean fibre length | 6 | 2.05 | 0.126 | 2 | 4.53 | 0.025 |
| 20% of tree height | ||||||
| Age at 1.5 mm | 6 | 2.14 | 0.112 | 2 | 6.49 | 0.008 |
| Age at 2.5 mm | 6 | 5.33 | 0.005 | 2 | 19.02 | 0.000 |
| Mean fibre length | 6 | 3.14 | 0.036 | 2 | 11.29 | 0.001 |
| 70% of tree height | ||||||
| Age at 1.5 mm | 6 | 0.47 | 0.817 | 2 | 0.38 | 0.690 |
| Age at 2.5 mm | 6 | 0.62 | 0.711 | 2 | 0.65 | 0.536 |
| Mean fibre length | 6 | 2.72 | 0.058 | 2 | 5.38 | 0.015 |
| Table 4. Number of year rings and mean ring width within different fibre length classes at breast height (BRH) and 20% of total tree height of the treatments examined. Different letters indicate significant differences. 300L = thinning leaving the 300 largest trees ha–1; 300S = thinning leaving the 300 smallest trees ha–1; P600 = PCT to 600 stems ha–1 at a dominant height of 5m; NT = no thinning; NP = no PCT; 3M = 3 m spacing; 10M = 10 m spacing. | ||||||
| Treatment | 0.3–1.5 mm | 1.5–2.5 mm | > 2.5 mm | |||
| No of years | Ring width (mm year–1) | No of years | Ring width (mm year–1) | Ring width (mm year–1) | ||
| BRH | ||||||
| 300L | 5.16ab | 2.98a | 14.30a | 3.21a | 1.60a | |
| 300S | 5.35ab | 2.48a | 15.20a | 1.99a | 1.53ab | |
| P600 | 4.47b | 2.62a | 12.59a | 2.99a | 1.51ab | |
| NT | 5.40ab | 2.29a | 11.68a | 2.22a | 1.05ab | |
| NP | 7.08ab | 3.21a | 13.40a | 1.74a | 0.73b | |
| 3M | 6.75ab | 3.16a | 14.96a | 2.98a | 1.30ab | |
| 10M | 10.81a | 3.63a | 17.38a | 2.86a | 1.40ab | |
| 20% | ||||||
| 300L | 4.42a | 3.56a | 11.35ab | 3.64a | 1.80a | |
| 300S | 4.03a | 3.78a | 11.36ab | 2.22b | 1.44ab | |
| P600 | 5.45a | 3.01a | 11.62ab | 3.23ab | 1.42ab | |
| NT | 4.60a | 3.56a | 8.28b | 2.45ab | 1.14ab | |
| NP | 4.58a | 3.46a | 8.85b | 2.46ab | 0.82b | |
| 3M | 7.05a | 3.72a | 14.64ab | 2.83ab | 1.26ab | |
| 10M | 6.82a | 3.57a | 15.72a | 3.75a | 1.46ab | |
At both breast height and 20%, the longest time taken to produce mature fibres occurred in trees managed in the Sparse regime. At breast height, it took, on average, 24.9 years before 2.5 mm long fibres were produced in the Sparse regime while it took 18.8 and 19.0 years for Dense and Dense/Sparse respectively to produce mature fibres. These differences were not statistically significant. However, the time taken before 1.5 mm long fibres were produced differed statistically between the Dense/Sparse and Sparse regimes (Table 5).
At 20% of the total tree height, the Sparse regime differed statistically in a negative way from the two other regimes in terms of time before producing both 1.5 and 2.5 mm long fibres. At that height, the Dense regime reached mature fibre length in 13.2 years which was the significantly shortest time. Corresponding figures for the other regimes were 16.1 (Dense/Sparse) and 22.1 (Sparse) years (Table 5).
| Table 5. Number of year rings and mean ring width within different fibre length classes at breast height (BRH) and 20% of total tree height of the regime types examined. Different letter indicate significant differences. Dense/Sparse = a high initial stand density followed by high intensity pre-commercial thinning/commercial thinning, Dense = a continuous high stand density and Sparse = a continuous low stand density. | ||||||
| Regime type | 0.3–1.5 mm | 1.5–2.5 mm | > 2.5 mm | |||
| No of years | Ring width (mm year–1) | No of years | Ring width (mm year–1) | Ring width (mm year–1) | ||
| BRH | ||||||
| Dense | 6.24ab | 2.75a | 12.54a | 1.98b | 0.89b | |
| Dense/Sparse | 4.99b | 2.69a | 14.03a | 2.73ab | 1.54a | |
| Sparse | 8.78a | 3.40a | 16.17a | 2.92a | 1.35a | |
| 20% | ||||||
| Dense | 4.59b | 3.51a | 8.56c | 2.46a | 0.98b | |
| Dense/Sparse | 4.63b | 3.45a | 11.44b | 3.03a | 1.55a | |
| Sparse | 6.94a | 3.64a | 15.18a | 2.29a | 1.36a | |
3.2 Mean fibre length, and proportions of fibre length classes
Mean fibre length was found to be influenced by both treatment and regime type (Table 3). The greatest mean fibre length was found below the green crown in all regimes. The Sparse regime showed the lowest mean fibre length at all sample heights. However, there were large between-tree variations. Compared to the Dense regime, the fibre length in the Dense/Sparse regime decreased with increases in tree height (Fig. 1).
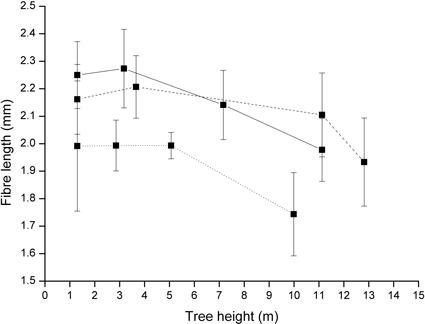
Fig. 1. Mean fibre length at the sample heights of the management regimes examined. Solid line represents Dense/Sparse (= a high initial stand density followed by high intensity pre-commercial thinning/commercial thinning), dashed line represents Dense (= a continuous high stand stem density) and dotted line denote Sparse (= a continuous low stand stem density); bars denote standard deviations.
The treatments 300L, P600 and 300S had the lowest proportions of wood containing fibres shorter than 1.5 mm (Fig. 2). The highest proportion of the JW fibre length class was found in trees managed in the Sparse regime, although differences were generally small and not statistically significant. Both regimes with initial high stand stem densities produced a significantly higher proportion of MW (fibres longer than 2.5 mm) compared with the trees growing in the Sparse regime. This difference was rather consistent for every treatment included in the regimes (Fig. 2). The difference was apparent in every stem section analyzed (Fig. 3). Consequently, the Sparse regime displayed high proportions of TW. Mean tree volume and volume by fibre class at different tree heights for the regimes is further shown in Fig. 4.
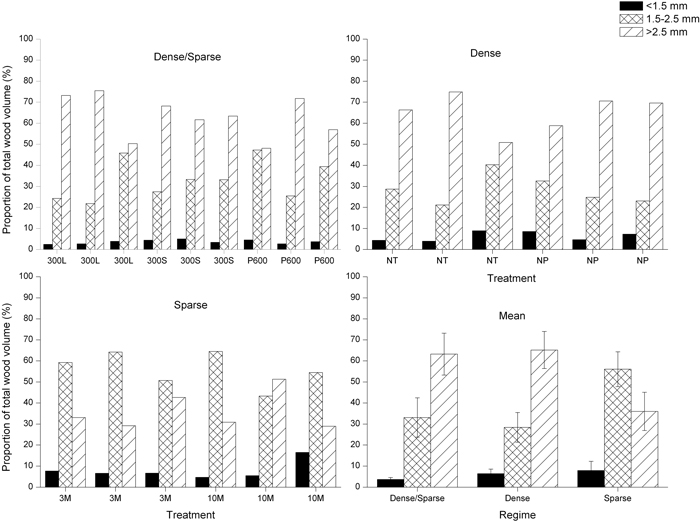
Fig. 2. Volume proportions of different fibre length intervals in all trees examined in each regime and mean values for the different principal management regimes (Dense/Sparse = a high initial stand density followed by high intensity pre-commercial thinning/commercial thinning, Dense = a continuous high stand stem density and Sparse = a continuous low stand stem density); bars denote standard deviations. 300L = thinning leaving the 300 largest trees ha–1; 300S = thinning leaving the 300 smallest trees ha–1; P600 = PCT to 600 stems ha–1 at a dominant height of 5 m; NT = no thinning; NP = no PCT; 3M = 3 m spacing; 10M = 10 m spacing.
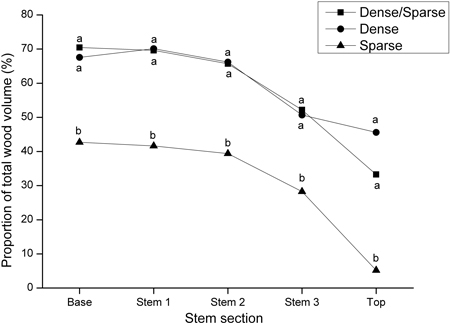
Fig. 3. Proportions of wood with fibres longer than 2.5 mm in relation to the total wood volume in five investigated stem sections from the base to the top of sample trees (Base = 0 to 1.3 m; Stem 1 = 1.3 m to 20% of total tree height; Stem 2 = 20% of total tree height to the height of the living crown; Stem 3 = height of the living crown to 70% of total tree height; Top = 70% of total tree height to 100% of total tree height). Different letters indicate significant differences between silvicultural regimes (Dense/Sparse = a high initial stand density followed by high intensity pre-commercial thinning/commercial thinning, Dense = a continuous high stand stem density and Sparse = a continuous low stand stem density). Squares denote Dense/Sparse regime, circles represent Dense regime and triangles show Sparse regime.
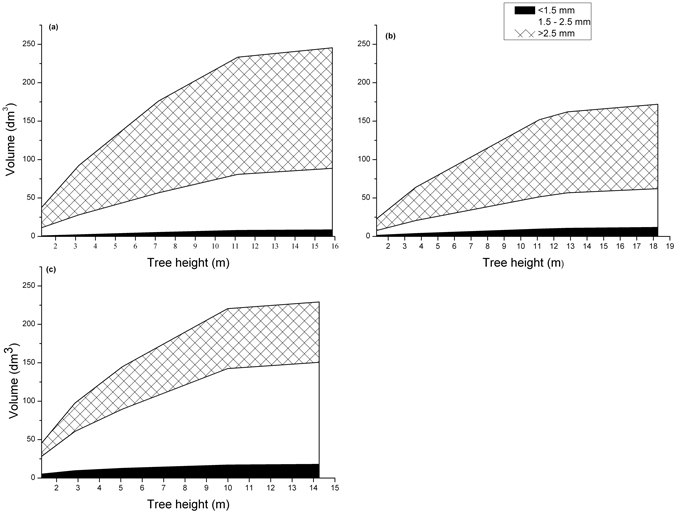
Fig. 4. Development of mean tree volume and wood volume by fibre length class with tree height in silvicultural management regimes with a high initial stand density followed by high intensity pre-commercial thinning/commercial thinning (a), a continuous high stand stem density (b) and a continuous low stand stem density (c).
4 Discussion
4.1 Effects of stand density on the proportion of juvenile wood, fibre length and associated quality traits
The results indicate that the choice of silvicultural regime can be a significant parameter that influences the volume of MW formed during one rotation period. The proportion of mature wood was clearly lower in the Sparse regime compared to both regimes with initially high stand stem densities. This indicates that the early environmental conditions that a tree faces are of great importance to the wood quality output produced over a rotation period. It also seems as if, in this respect, the negative effects of an initial sparse stand density are maintained throughout the major part of the life cycle of the trees. In the Dense/Sparse regime the proportion of JW amounted to 2.5–5% whereas corresponding proportions in Dense and Sparse regimes were 4–9 and 4.5–16% respectively. Therefore, the results indicate that the proportion of JW can be kept at a low level if initially slow grown trees are retained after high intensity thinning. However, mean fibre length in the upper part of the stem seems to be somewhat shorter in such a regime, compared to a non-thinning regime. This conclusion is further supported by Jaakkola et al. (2005), as they showed that Norway spruce (Picea abies) fibres were 4–9% shorter after intensive thinning than after low-intensity thinning. By applying appropriate management regimes, high quality wood and fibres for specific end-uses could be produced. Thereby, less focus on high volumes would be needed within the forest industry. However, whether it would be profitable to adjust silvicultural regimes according to expectations on fibre lengths require further research.
The shortest mean fibre length was found in trees growing in a regime where the distances between trees were kept relatively high throughout the rotation period. Short fibres have also previously been noted in widely-spaced plantations (Yang and Hazenberg 1994). Fibre morphology is an important parameter for paper quality properties as it influences strength and optical properties (Barefoot et al. 1966; Seth and Page 1988). According to Cao et al. (2008), the pulpwood price can be expected to increase by 2% for each 0.5 mm increase in fibre length. Therefore, regimes with initial low stand densities may not be optimal if the overall aim is to produce pulp and paper. Moreover, short fibre lengths have been associated with a large microfibril angle (Megraw 1985) which, in turn, contributes to low strength and stiffness in wood. According to results presented by Auty et al. (2013), the microfibril angle of Scots pines (with a ring width of 4 mm) decreases from about 25° to 20° within the JW zone. This decrease indicates that the modulus of elasticity (MOE) is about 23% (Cave 1968) higher in MW compared to JW. Similarly, a high initial stand density seems to have positive effects on the occurrence of knots and tapering (Persson 1977), branch diameter growth (Johansson 1992; Varmola and Salminen 2004) and relative density (Yang and Hazenberg 1994; Yang 2002). Thus, young, dense forests seem to be able to produce trees with several commercially preferred wood quality traits.
4.2 Transition ages
Treatment results indicated that it took longer for the trees that initially faced little competition to form fibres of mature length. At breast height, the mature state was reached within 17–28 years, which corresponds to previous findings for Scots pine. Using density as the defining parameter, Mutz et al. (2004) found that the transition to mature wood occurred between 18 and 27 years, whereas Sauter et al. (1999) reported the age to be 22 years. Fries et al. (2003) measured fibre length and concluded that the transition age was 16. By comparison of conceptual regimes, we found the stated hypothesis to be true. On average, trees within the Sparse regime formed mature wood five years later than the other regimes at breast height and six (compared to Dense/Sparse regime) to nine (compared to Dense regime) years later at 20% of total tree height. Similar results about the effects of the initial spacing have been presented for Norway spruce (Kucera 1994). Also, results presented by Eriksson et al. (2006) suggest that the initial growing conditions of a tree can regulate the timing of the transition between JW and MW. Clark and Saucier (1991) and Yang (1994) did not, however, find that the transition age between JW and MW was affected by the initial spacing, a finding which was further supported by Alteyrac et al. (2006). In the current study, transition ages decreased between breast height and 20% of total tree height. There was a significant difference in the age at which the fibre length reached 2.5 mm between Dense and Dense/Sparse regimes at 20% of the total tree height, indicating that thinning/PCT treatments have had some effect at that height.
Briggs and Smith (1986) concluded that with a large tree crown, the juvenile period is prolonged and more JW will be produced. In Norway spruce, Kucera (1994) found that the transition zone coincided with the culmination of current annual height increment. Furthermore, Bendtsen and Senft (1986) described how planted stands were likely to display higher relative amounts of JW than naturally regenerated stands would. Clark and Saucier (1991) found that the JW diameter increased with initial spacing and Yang (1994) has reported a juvenile zone whose area is about four times greater in 3.6 × 3.6 m spaced stands than that in 1.8 × 1.8 m spaced stands for white spruce and black spruce. Zobel and van Buijtenen (1989) reported that a decreased JW diameter in tight spacings does not necessarily result in a smaller percentage of JW. Instead, they found an increased proportion of JW in such plantations due to a low total tree basal area.
4.3 Study design
As well as traditional comparisons of treatments, this study was designed to compare trees with different silvicultural backgrounds, including different detailed treatments. To reduce the number of laborious analyses, the sampling procedure was restricted to three sample trees from each treatment and to four sampling heights at each tree. The procedure of randomly selecting sample trees will most likely have led to larger variations within regimes than if, for instance, trees mirroring the average tree within the stand had been consistently selected. However, variations between trees within regimes were still much smaller than the variations between regimes and this study therefore demonstrates the potential of regulating wood quality by the choice of silvicultural regime. Moreover, the results demonstrate that the initial environment affects the wood quality output for most of the rotation period. Using a similar approach, Eriksson et al. (2006) also managed to demonstrate that management regimes can be used as a tool to regulate structural characteristics and mechanical properties within trees. In the current study, the study sites were located in the same geographical area and they were all of comparable site quality. Therefore, they would all have experienced similar abiotic factors meaning that the silvicultural background might be an explanatory parameter for any differences in fibre length development. Mansfield et al. (2009) previously showed that site had no significant effect on the transition age as determined by fibre length. However, genetic variations between tree individuals can be of influence (Abdel-Gadir and Krahmer 1993; Gapare et al. 2006).
The fibre length limits used to describe the different wood types was based on findings for Norway spruce. According to the difference in fibre length between Norway spruce and Scots pine showed by Duchesne et al. (1997), the proportion of MW might have been slightly underestimated in the current study.
4.4 Conclusions
The results of this study indicate that wood properties and quality to some degree can be regulated by silvicultural long-term strategies. Initial stand stem densities were found to influence the juvenile to mature transition age. Management regimes including low initial stand stem numbers are therefore likely to result in short mean fibre length and display relatively low proportions of mature wood. Moreover, it was found that the proportion of juvenile wood could be regulated by high intensity thinning. Importantly, the early environmental conditions that a tree faces have a large impact on the overall wood quality throughout most of its life cycle. Thus, this study highlights that long-term strategies implemented at juvenile stand ages can be important tools in order to produce wood raw materials suited for specific end-uses.
Acknowledgements
This study was founded by the Swedish Energy Agency and by the Kempe and Brattås foundations. We are grateful to Sees editing for editorial assistance, to Dr. Anders Muszta for statistical assistance, to Dr. Thomas Ulvcrona for valuable comments regarding the manuscript, and to Hans-Göran Nilsson of the Vindeln Experimental Field Station for supplying us with the equipment needed.
References
Abdel-Gadir A.Y., Krahmer R.L. (1993). Genetic variation in the age of demarcation between juvenile and mature wood in Douglas fir. Wood and Fiber Science 25: 384–394.
Alteyrac J., Cloutier A., Zhang S.Y. (2006). Characterization of juvenile wood to mature wood transition age in black spruce (Picea mariana (Mill.) B.S.P.) at different stand densities and sampling heights. Wood Science and Technology 40: 124–138. http://dx.doi.org/10.1007/s00226-005-0047-4.
Auty D., Gardiner B.A., Achim A., Moore J.R., Cameron A.D. (2013). Models for predicting microfibril angle variation in Scots pine. Annals of Forest Science 70: 209–218. http://dx.doi.org/10.1007/s13595-012-0248-6.
Barefoot A.C., Hitchings R.G., Ellwood E.L. (1966). Wood characteristics and kraft paper properties of four selected loblolly pines. Tappi Journal 49: 137–147.
Bendtsen B.A. (1978). Properties of wood from improved and intensively managed trees. Forest Products Journal 10: 61–72.
Bendtsen B.A., Senft J. (1986). Mechanical and anatomical properties in individual growth rings of plantation-grown eastern cottonwood and loblolly pine. Wood and Fiber Science 18: 23–38.
Boutelje J. (1968). Juvenile wood, with particular reference to northern spruce. Svensk pappers-tidning 17: 581–585.
Briggs D.G., Smith R. (1986). Effects of silvicultural practices on wood properties of conifers: a review. In: Oliver C., Hanley D., Johnson J. (eds.). Douglas fir: stand management for the future. University of Washington Press, Seattle, WA. p. 108–117.
Cao T., Valsta L., Härkönen, S, Saranpää P., Mäkelä A. (2008). Effects of thinning and fertilization on wood properties and economic returns for Norway spruce. Forest Ecology and Management 256: 1280–1289. http://dx.doi.org/10.1016/j.foreco.2008.06.025.
Cave I.D. (1968). The anisotropic elasticity of the plant cell wall. Wood Science and Technology 2: 268–278. http://dx.doi.org/10.1007/BF00350273.
Clark A., Saucier J.R. (1989). Influence of initial planting density, geographic location, and species on juvenile wood formation in southern pine. Forest Products Journal 7/8: 42–48.
Dinwoodie J.M. (2000). Timber: its nature and behaviour. 2 ed. E. & F.N. Spon, London. 257 p.
Duchesne I., Wilhelmsson L., Spångberg K. (1997). Effects of in-forest sorting of Norway spruce (Picea abies) and Scots pine (Pinus sylvestris) on wood and fibre properties. Canadian Journal of Forest Research 27: 790–795. http://dx.doi.org/10.1139/x97-040.
Eriksson D., Lindberg H., Bergsten U. (2006). Influence of silvicultural regime on wood structure characteristics and mechanical properties of clear wood in Pinus sylvestris. Silva Fennica 40: 743–762.
Fahlvik N., Ekö P.M & Pettersson N. (2005). Influence of precommercial thinning grade on branch diameter and crown ratio in Pinus sylvestris in southern Sweden. Scandinavian Journal of Forest Research 20: 243–251. http://dx.doi.org/10.1080/02827580510008266.
Franklin G. (1945). Preparations of thin sections of synthetic resins and wood-resin composites, a new macerating method for wood. Nature 155: 51–51. http://dx.doi.org/10.1038/155051a0.
Fries A., Ericsson T., Mörling T. (2003). Measuring relative fibre length in Scots pine by non-destructive wood sampling. Holzforschung 4: 400–406.
Gapare W.J., Wu H.X., Abarquez A. (2006). Genetic control of the time of transition from juvenile to mature wood in Pinus radiata D. Don. Annals of Forest Science 63: 871–878. http://dx.doi.org/10.1051/forest:2006070.
Hägglund B. (1974). Site index curves for Scots pine in Sweden. Swedish University of Agricultural Sciences, Department of Forest Yield Research, Report 31: 1–54.
Harris J.M. (1981). Wood quality of radiata pine. Appita 3: 211–215.
Jaakkola T., Mäkinen H., Sarén M-P., Saranpää P. (2005). Does thinning intensity affect the tracheid dimensions of Norway spruce? Canadian Journal of Forest Research 35: 2685–2697. http://dx.doi.org/10.1139/x05-182.
Johansson K. (1992). Effects of initial spacing on the stem and branch properties and graded quality of Picea abies (L.) Karst. Scandinavian Journal of Forest Research 7: 503–514. http://dx.doi.org/10.1080/02827589209382743.
Karlsson K., Ulvcrona T. (2010). Field experiment data available for studies of pre-commercial thinning. Swedish University of Agricultural Sciences, Unit for Field-based Forest Research, Vindeln Experimental Forests. Skog & Trä 2010:2: 1–46. ISBN 978-91-977896-3-9.
Kirk D.G., Breeman L.G., Zobel B.J. (1972). A pulping evaluation of juvenile loblolly pine. Tappi Journal 11: 1600–1604.
Kucera B. (1994). A hypothesis relating current annual height increment to juvenile wood formation in Norway spruce. Wood and Fiber Science 26: 152–167.
Mansfield S.D., Parish R., Di Lucca C.M., Goudie J., Kang, K-Y., Ott P. (2009). Revisiting the transition between juvenile and mature wood: a comparison of fibre length, microfibrill angle and relative wood density in lodgepole pine. Holzforschung 63: 449–456. http://dx.doi.org/10.1515/HF.2009.069.
Megraw R.A. (1985). Wood quality factors in loblolly pine. The influence of tree age, position in tree, and cultural practice on wood specific gravity, fiber length, and fibril angle. Tappi Press, Atlanta. 88 p.
Minitab Statistical Software Release 15 for Windows. (2007).
Mutz R., Guilley E., Sauter U.H., Nepveu G. (2004). Modelling juvenile-mature wood transition in Scots pine (Pinus sylvestris L.) using nonlinear mixed-effects models. Annals of Forest Science 61: 831–841. http://dx.doi.org/10.1051/forest:2004084.
Myers G.C. (2001). Thermomechanical pulping of loblolly pine juvenile wood. Wood and Fiber Science 1: 108–115.
Nilsson U., Agestam E., Ekö P-M., Elfving B., Fahlvik N., Johansson U., Karlsson K., Lundmark T., Wallentin C. (2010). Thinning of Scots pine and Norway spruce monocultures in Sweden – effects of different thinning programmes on stand level gross- and net stem volume production. Studia Forestalia Suecica 19: 1–46. ISBN 978-91-86197-76-6.
Panshin A.J., de Zeeuw C. (1980). Textbook of wood technology. 4 ed. McGraw-Hill Book Company. 722 p.
Pearson R.G., Gilmore R.C. (1971). Characterization of the strength of juvenile wood of loblolly pine (Pinus taeda L.). Forest Products Journal 1: 23–31.
Persson A. (1977). Quality development in young spacing trials with Scots pine. Swedish University of Agricultural Science, Department of Forest Yield Research, Report 45. 152 p.
Saranpää, P. (1994). Basic density, longitudinal shrinkage and tracheid length of juvenile wood of Picea abies (L.) Karst. Scandinavian Journal of Forest Research 9: 68–74. http://dx.doi.org/10.1080/02827589409382814.
Sauter U.H., Mutz R., Munro B.D. (1999). Determining juvenile-mature wood transition in Scots pine using latewood density. Wood and Fiber Science 31: 416–425.
Seth R.S., Page D.H. (1988). Fiber properties and tearing resistance. Tappi Journal 71: 103–107
Swedish Statistical Yearbook of Forestry. (2011). Swedish Forest Agency, Jönköping.
Ulvcrona T., Ulvcrona K.A. (2011). The effects of pre-commercial thinning and fertilization on characteristics of juvenile clearwood of Scots pine (Pinus sylvestris L.). Forestry 3: 207–219. http://dx.doi.org/10.1093/forestry/cpr007.
Uprichard J.M., Lloyd J.A. (1980). Influence of tree age on the chemical composition of radiata pine. New Zealand Journal of Forestry Science 3: 551–557.
Varmola M., Salminen H. (2004). Timing and intensity of precommercial thinning in Pinus sylvestris stands. Scandinavian Journal of Forest Research: 19: 142–151. http://dx.doi.org/10.1080/02827580310019545.
Yang K.C. (1994). Impact of spacing on width and basal area of juvenile and mature wood in Picea mariana and Picea glauca. Wood and Fiber Science 26: 479–488.
Yang K.C. (2002). Impact of spacing on juvenile wood and mature wood properties of white spruce (Picea glauca). Taiwan Journal of Forest Science 17: 13–29.
Yang K.C., Benson C.A., Wong J.K. (1986). Distribution of juvenile wood in two stems of Larix laricina. Canadian Journal of Forest Research 16: 1041–1049. http://dx.doi.org/10.1139/x86-181.
Yang K.C., Hazenberg G. (1994). Impact of spacing on tracheid length, relative density, and growth rate of juvenile wood and mature wood in Picea mariana. Canadian Journal of Forest Research 24: 996–1007. http://dx.doi.org/10.1139/x94-130.
Zhu J., Nakano T., Hirakawa Y. (2000). Effects of radial growth rate on selected indices for juvenile and mature wood of the Japanese larch. Journal of Wood Science 46: 417–422. http://dx.doi.org/10.1007/BF00765798.
Zobel B.J., Sprague J.R. (1998). Juvenile wood in forest trees. Springer-Verlag, Berlin, Germany. 300 p. http://dx.doi.org/10.1007/978-3-642-72126-7.
Zobel B.J., van Buijtenen J.P. (1989). Wood variation: its cause and control. Springer-Verlag, Berlin, Germany. 363 p. http://dx.doi.org/10.1007/978-3-642-74069-5.
Total of 48 references
