Claudie-Maude Canuel,
Nelson Thiffault  ,
Michael K. Hoepting,
James C.G. Farrell
,
Michael K. Hoepting,
James C.G. Farrell
Legacy effects of precommercial thinning on the natural regeneration of next rotation balsam fir stands in eastern Canada
Canuel C.-M., Thiffault N., Hoepting M. K., Farrell J. C.G. (2019). Legacy effects of precommercial thinning on the natural regeneration of next rotation balsam fir stands in eastern Canada. Silva Fennica vol. 53 no. 4 article id 10209. https://doi.org/10.14214/sf.10209
Highlights
- We investigated the potential legacy effects of precommercial thinning in next rotation, dense natural balsam fir stands
- Precommercial thinning had few legacy effects on next rotation stands and should not impair their regeneration
- Balsam fir dominated the regeneration layer. Other tree species were almost absent.
Abstract
The Green River precommercial thinning (PCT) trial was established between 1959–1961 in New Brunswick (Canada) within natural balsam fir (Abies balsamea (L.) Mill.)-dominated stands. Three silviculture scenarios differing only by the increasing nominal spacings of PCT treatments (1.2 m, 1.8 m, 2.4 m) were compared to an unthinned control within randomized replicates that were clearcut harvested in 2008 and treated with herbicide in 2011. During the fourth post-harvest growing season, we assessed regeneration, competing vegetation and coarse woody debris (CWD; differentiated between large woody debris and slash) to assess the legacy effects of PCT on regeneration of next rotation stands. Our results confirmed that silviculture scenarios including PCT significantly increased conifer stocking in treated plots compared to control conditions, but only in the 1.8 m nominal spacing. Considering that treated and untreated stands were fully stocked, we conclude that PCT using the spacing gradient tested has no legacy effect on the regeneration of next rotation natural balsam fir stands. Given the known sensitivity of balsam fir to future climate conditions in this region, we suggest that future treatments should promote tree species diversity to support ecosystem resilience to climate change by favouring more warm-adapted species, such as some hardwoods.
Keywords
silviculture;
Abies balsamea;
PCT;
conifer;
density management
- Canuel, Faculté de foresterie, géographie et géomatique, Université Laval, Québec, QC G1V 0A6, Canada; Canadian Wood Fibre Centre, Natural Resources Canada, 1055 du P.E.P.S., P.O. Box 10380, Sainte-Foy Stn., Québec, QC G1V 4C7, Canada E-mail claudie-maude.canuel.1@ulaval.ca
-
Thiffault,
Canadian Wood Fibre Centre, Natural Resources Canada, 1055 du P.E.P.S., P.O. Box 10380, Sainte-Foy Stn., Québec, QC G1V 4C7, Canada
 http://orcid.org/0000-0003-2017-6890
E-mail
nelson.thiffault@canada.ca
http://orcid.org/0000-0003-2017-6890
E-mail
nelson.thiffault@canada.ca
- Hoepting, Canadian Wood Fibre Centre, Natural Resources Canada, 1219 Queen St. E., Sault Ste. Marie, ON P6A 2E5, Canada E-mail michael.hoepting@canada.ca
- Farrell, Canadian Wood Fibre Centre, Natural Resources Canada, 1350 Regent Street, P.O. Box 4000, Fredericton, NB E3B 5P7, Canada E-mail jamescg.farrell@canada.ca
Received 12 June 2019 Accepted 11 October 2019 Published 17 October 2019
Views 73344
Available at https://doi.org/10.14214/sf.10209 | Download PDF

1 Introduction
Precommercial thinning (PCT) is a silviculture treatment commonly used in young stands characterized by high stem densities originating from dense regeneration. The main objective of PCT is to lower stand density to redistribute site resources and growing space of residual trees, so that competition does not reduce individual tree growth and stand yield (Nyland et al. 2016). Indeed, PCT has been shown to significantly increase soil temperature and decomposition rates resulting in short-term nutrient availability for residuals trees (Piene 1978; Thibodeau et al. 2000). These effects, combined with reduced competition for environmental resources (Piene 1978; Pothier and Margolis 1991), result in enhanced diameter growth and individual stem volume (Pothier 2002; Pitt and Lanteigne 2008; Zhang et al. 2009), although they rarely increase overall stand productivity (Zeide 2001; Pitt and Lanteigne 2008). Precommercial thinning has been shown to optimize timber production by generating more sawlog volume per tree (Pitt et al. 2013b). The increased growth rate of individual stems can reduce rotation length by producing larger trees quicker, and lower harvesting costs through increasing harvesting efficiencies (Pitt and Lanteigne 2008; Zhang et al. 2009; Plamondon and Pitt 2013; Pitt et al. 2013b). Precommercial thinning is also an opportunity for controlling stand composition (Prévost and Gauthier 2012) and selecting the most vigorous crop trees.
Juvenile balsam fir (Abies balsamea (L.) Mill.) stands of eastern Canada are particularly favorable to PCT (Gauthier and Tremblay 2018). They usually have dense and abundant regeneration (Ruel 1989), so the intraspecific competition is high (Frank 1990). Indeed, balsam fir trees older than 20 y produce high amount of seeds that are mainly dispersed within less than 60 m (Frank 1990; Ontario Ministry of Natural Resources 2000). As a shade tolerant species (Humbert et al. 2007), balsam fir builds large seedling banks in the understory of mature stands that survive many decades (Frank 1990). Advanced balsam fir regeneration reacts positively to increased light levels following canopy removal by harvesting or natural disturbances (Côté and Bélanger 1991; Pothier and Margolis 1991; Côté 2013). In southeastern Canada, regenerating balsam fir stands may contain more than 25 000 seedlings ha–1 (Baldwin 1977; Côté and Bélanger 1991; Brissette 1996). From 2007 to 2017, approximately 85 000 ha have been treated annually by PCT in eastern Canada, mainly in Quebec, New Brunswick, Ontario and Nova Scotia (Canadian Council of Forest Ministers 2019).
While short-term (e.g. 10 y following treatment) and long-term (e.g. rotation age) effects of PCT are well documented, to our knowledge, potential legacy effects of PCT on the regeneration of next rotation stands has rarely been investigated. Because of its impacts on stand structure, density, crown development and light regime in the understory, PCT has the potential to influence vegetation, woody debris and regeneration. Precommercial thinning can indeed modify the composition and abundance of understory vegetation (Lindgren et al. 2006), although effects can be modest in mature stands and vary over time because of residual tree growth (Hale 2003; Cole et al. 2008, 2010) and self-thinning (Pitt and Lanteigne 2008). Moreover, the increasing presence of coarse woody debris (CWD) post-harvest (Fraver et al. 2002), which can be affected by initial stocking (Sturtevant et al. 1997) and wood utilization, may affect regeneration as CWD create physical barriers that limit access to space and light, thereby reducing soil temperature and its fluctuation (Trottier-Picard et al. 2014).
Balsam fir is a cold-adapted boreal species that is at high-risk under the predicted future climate conditions, particularly in the Acadian Region of Canada (Bourque and Hassan 2008; Taylor et al. 2017). Additionally, there are increasing concerns that the cumulative effects of forest management activities are posing a risk to the sustainability of both Canada’s forests and forest industry, especially with the added pressure of climate change (Gauthier et al. 2014). Hence, there is a need to document the effects of PCT on regeneration characteristics beyond a single rotation to evaluate if and how this common treatment is affecting the sustainability of forest activities.
In this context, our objective was to evaluate the legacy effects of silviculture scenarios that include PCT treatments of varying intensity on natural regeneration in harvested balsam fir stands, as well as on CWD cover and competing vegetation cover. We hypothesized that PCT affects the density (stems ha–1) and growth of regenerating conifers during the subsequent rotation, through its legacy impacts on stand characteristics, including production of CWD. We predicted that the stocking, density and growth of balsam fir in regenerating stands would increase according to PCT intensity applied to the previous stand.
2 Materials and methods
2.1 Study area
The study was conducted in northwestern New Brunswick, Canada (47°47´N, 68°19´W), in the 100 000 ha Green River watershed (Baskerville et al. 1960; Pitt et al. 2013a) (Fig. 1). The study area is comprised within the Northwestern Plateau geomorphological unit (Loucks 1962). The rough topography shows a peneplain of 300 to 450 m elevation a.s.l., with steep slopes and many streams. The Paleozoic slate and argillite bedrock forms shallow and stony soils. Climate is continental, with abundant precipitation and large fluctuations in temperature (Loucks 1962). Based on the nearest meteorological station (Edmundston, New Brunswick, 47°22´N, 68°19´W), the study area receives an average precipitation of 1011 mm year–1, of which almost half falls between May and October (Environment Canada 2010). The mean annual temperature is 3.6 °C. The study area is within the Gaspésie section of the Boreal Forest ecological region defined by Rowe (1972), and within the Highlands ecoregion defined by the Department of Natural Resources of New Brunswick (2007). The most common tree species include balsam fir, white spruce (Picea glauca (Moench) Voss), black spruce (Picea mariana (Mill.) Britton, Sterns & Poggenb.), sugar maple (Acer saccharum Marshall), red maple (Acer rubrum L.), white birch (Betula papyrifera Marshall), yellow birch (Betula alleghaniensis Britton), northern white cedar (Thuja occidentalis L.), beech (Fagus grandifolia Ehrh.) and poplar (Populus spp.) (Baskerville et al. 1960).
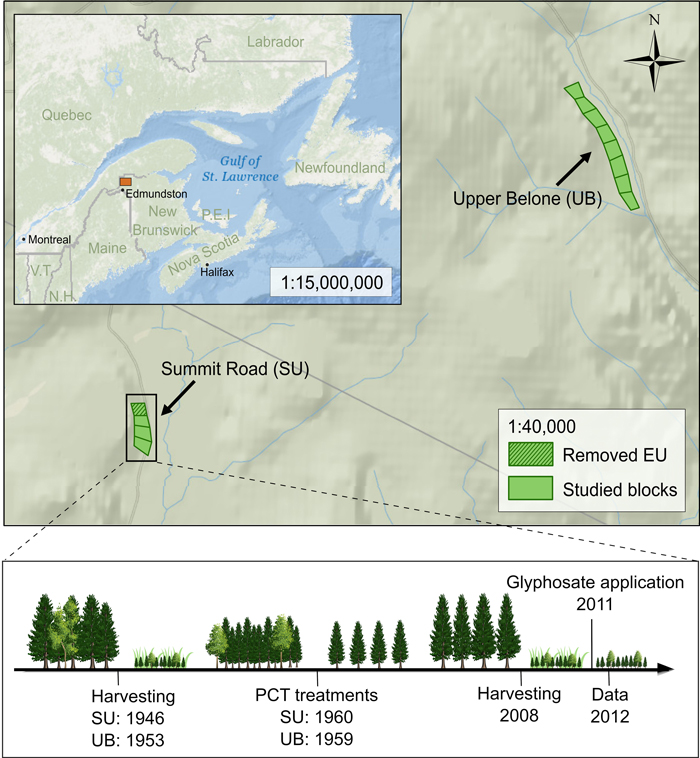
Fig. 1. Replicate blocks and sequence of treatments of the Green River precommercial thinning study within balsam fir stands in northwestern New Brunswick (Canada). The studied blocks contained four experimental units (EU; 1–2 ha) that were treated with silviculture scenarios including one of three increasing precommercial thinning (PCT) intensity (1.2 m nominal spacing, 1.8 m nominal spacing, 2.4 m nominal spacing) or an unthinned control. The timeline provides the date for the silvicultural treatments applied. Aerial glyphosate application was of 1.62 kg ha–1 of active ingredient. Two blocks are side-by-side at UB. Note that the 1.8 m nominal spacing treatment at SU (hatched EU in the figure) was excluded from the analyses due to major blowdown from the adjacent stand, hence resulting in an incomplete randomized block design. The chronosequence was designed using resources from Freepik.com.
2.2 Experimental design and treatments
The study area was clearcut harvested between 1946 and 1955 (Fig. 1). Balsam fir dominated the previous stands, with minor components of red spruce (Picea rubens Sarg.) and white birch. Between 1959 and 1961, six replicated blocks that each contained four main plots of 1–2 ha each were established (Pitt and Lanteigne 2008). Hence, the initial study design comprised a total of 24 experimental units (EU) (6 blocks × 4 main plots). The plots were rapidly dominated by dense natural conifer regeneration. Six to fourteen years after harvesting, each main plot (EU) of each block randomly received one of four PCT treatments of increasing intensity: control (no thinning), 1.2 m nominal spacing, 1.8 m nominal spacing, or 2.4 m nominal spacing. In 2008, three of the original 6 blocks were clearcut harvested again to evaluate the effect of PCT on volume production, product quantity and quality (Bicho et al. 2013; Duchesne et al. 2013; Pitt et al. 2013b), economic return at rotation age (Pitt et al. 2013c) and overall harvesting efficiency (Plamondon and Pitt 2013). Between mid-August to mid-September 2011, the three harvested blocks (12 EU) were treated with an aerial herbicide application (4.5 L ha–1 of glyphosate formulation at 360 g L–1, hence 1.62 kg ha–1 of active ingredient), as part of standard juvenile stand tending in New Brunswick, to reduce competition from shrubs and deciduous trees (Forest Management Branch 2004; Department of Natural Resources of New Brunswick 2008). One of the 12 EU of the three harvested blocks suffered from partial blowdown due to an adjacent cutover prior 2004 and was excluded from the experimental design.
2.3 Assessment of natural regeneration in subsequent rotation stands
Our experiment focused on the silviculture scenarios in three blocks that were harvested in 2008 using an incomplete randomized block design of 11 experimental units (Fig. 1). Our study thus compared three silviculture scenarios differing only by the increasing nominal spacings of PCT treatments that were applied (1.2 m, 1.8 m, 2.4 m) plus an unthinned control. In July and September 2012, we collected data on regeneration in the individual treatment plots, 52 or 53 years after the initial PCT treatment (depending on block), during the fourth post-harvest growing season (Fig. 1). To do so, we established a total of 396 regeneration sampling plots (2 m × 2 m; equally distributed among the 11 EU (36 plots × 11 EU). Fourteen regeneration sampling plots were later removed from the database as they were not sprayed with the herbicide. In each EU, plots were centered along two parallel 68-m long transects, positioned 20 m apart (Fig. 2). We located the first transect at a minimal distance of 30 m from main roads and 20 m from the plot side. The second transect was also located at 20 m from plot side. Along each transect, we established 18 regeneration sampling plots with plot centers 4 m apart (2 transects × 18 plots). If a treatment plot was not wide enough (<108 m) to include a whole transect of 18 regeneration sampling plots, we added additional transects and distributed sampling plots along them.
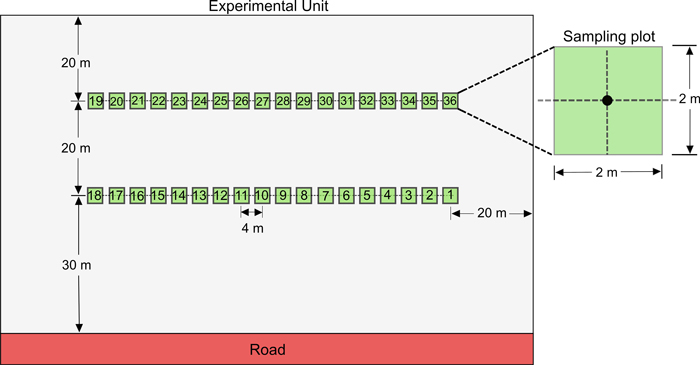
Fig. 2. Schematic representation of one of the 11 experimental units (EU) established within the three blocks. We established a total of 36 sampling plots within each EU. Sampling plot centers were spaced 4 m apart along two parallel 68-m long transects located 20 m apart. The minimal distances were 30 m and 20 m from main roads and EU limits, respectively.
To assess stocking of commercial species (conifers, other commercial species), we noted presence or absence of seedlings within each regeneration sampling plot. Then, we selected the spruce or balsam fir seedling (>1 y) with the best form and vigor (Ruel et al. 2000) and measured its height (cm), basal diameter 5 cm above ground line (mm), maximum crown diameter (CDmax; cm) and crown diameter perpendicular to the maximum axis (CDmin; cm). We used seedling crown data to calculate the crown area (CA; m2).
We counted the remaining conifer seedlings (spruce, balsam fir; >1 y) according to species within each sampling plot. We also collectively counted stems of other commercial species in a separate class. We then calculated the average stem density per hectare (balsam fir, spruce, other commercial species).
We visually assessed the percent of the sample plots covered with slash piles (made of several stems of woody debris), large woody debris including stumps (>5 cm diameter) and competing vegetation without considering conifer overtopping. We calculated percent cover of coarse woody debris by adding the percent cover of slash piles and the percent cover of large woody debris.
2.4 Statistical analyses
The main effects of silviculture scenarios differing only in PCT treatments on subsequent rotation regenerating stands were evaluated using linear mixed-effect models based on the experimental design (Figs. 1 and 2) with the function lme of package nlme (Pinheiro et al. 2018) in R 3.5.1 (R Core Team 2018); block and interactions involving block were considered as random effects, while PCT (3 levels and an unthinned control) was considered as a fixed effect treatment. We conducted analyses on seedling variables (height, basal diameter and crown area). We also tested treatment effect on competition (competing vegetation cover) and debris (slash cover and large woody debris cover), stem density (balsam fir density, spruce density, conifer density and other commercial species density) and stocking (conifer stocking and other commercial species stocking). We verified normality and homoscedasticity using standard graphical approaches (function plot) on residuals and using both graphical approaches (functions qqnorm and qqline) and Bartlett tests on raw data. We then applied inverse sine or square root transformations when necessary (Steel et al. 1997). We used α = 0.05 as threshold for significance for all analyses. When necessary, we proceeded with multiple comparisons for linear mixed effects models using Tukey’s tests with adjusted p-values with function glht of the multcomp package (Hothorn et al. 2019).
3 Results
3.1 Conifer seedling characteristics, competition and debris
Silviculture scenarios including PCT in the previous rotation did not have any significant effect on seedling height, basal diameter or crown area in regenerating stands (Table 1, Fig. 3). Moreover, four years following cut-to-length clearcut harvesting, there were no legacy effects of PCT intensity on the abundance of competing vegetation (competing vegetation cover) or the abundance of residual harvesting coarse woody debris (slash cover, large woody debris cover, total debris cover) (Table 1, Fig. 4).
| Table 1. ANOVA summary for conifer seedling characteristics, competition and debris, stem density and stocking in next rotation balsam fir stands as influenced by different intensities of precommercial thinning in previous stands. | |||
| Variable | F value | p-value | |
| Conifer seedling characteristics | Stem height (cm) ** | 1.182 | 0.405 |
| Stem basal diameter (mm) ** | 3.301 | 0.116 | |
| Crown area (cm2) ** | 0.962 | 0.479 | |
| Competition and debris | Slash cover (%) * | 2.490 | 0.175 |
| Large woody debris cover (%) * | 1.770 | 0.269 | |
| Total debris cover (%) * | 2.033 | 0.228 | |
| Competing vegetation cover (%) * | 0.387 | 0.768 | |
| Stem density | Balsam fir density (stems/ha) ** | 1.285 | 0.375 |
| Spruce density (stems/ha) ** | 0.089 | 0.963 | |
| Conifer density (stems/ha) ** | 1.188 | 0.403 | |
| Other species density (stems/ha) ** | 0.517 | 0.689 | |
| Stocking | Conifer stocking (%) * | 5.969 | 0.042 |
| Other species stocking (%) * | 0.401 | 0.759 | |
| For all analyses, degrees of freedom were 3 and 5 at numerator and denominator, respectively. We performed analyses on transformed data: * inverse sine transformation; ** square root transformation. We used p = 0.05 as a threshold for significance. | |||
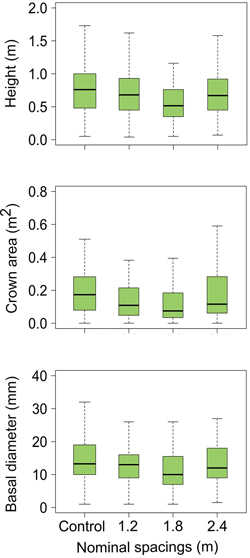
Fig. 3. Estimated values (lower hinge = within 1.5 times interquartile range below first quartile, lower end of box = first quartile, middle of box = median, upper end of box = third quartile, upper hinge = within 1.5 times interquartile range above third quartile) for conifer seedling characteristics in next rotation balsam fir stands as influenced by different intensities of precommercial thinning in previous stands.
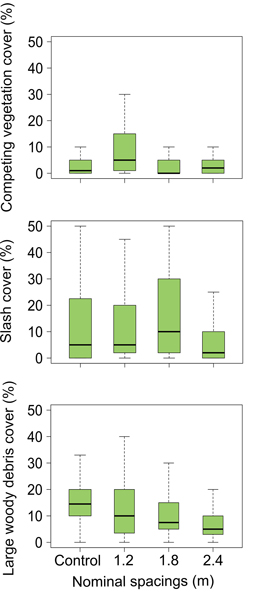
Fig. 4. Estimated values (lower hinge = within 1.5 times interquartile range below first quartile, lower end of box = first quartile, middle of box = median, upper end of box = third quartile, upper hinge = within 1.5 times interquartile range above third quartile) for competition and debris in next rotation balsam fir stands as influenced by different intensities of precommercial thinning in previous stands. Large woody debris include stumps and debris > 5 cm in diameter. Slash include piles made of several stems of woody debris.
3.2 Seedling density and stocking
Silviculture scenarios including PCT in the previous rotation did not have any significant effect on seedling density for all species (balsam fir, spruce) or species group (conifer, other species) of the next rotation stand (Table 1, Fig. 5). Other species stocking was equivalent for both treated and untreated stands, but conifer stocking appears to vary depending on PCT intensity (Table 1). Conifer stocking was higher in thinned stands originating from the 1.8 m nominal spacing than both untreated stands and stands originating from the 1.2 m nominal spacing (Fig. 6). As a general trend, balsam fir seedlings were considerably more abundant than spruce seedlings and other species in both treated and untreated plots (average proportion = 96% balsam fir, 4% spruce, <1% other species). Other species density was low (median = 0stems ha–1) in every plot.
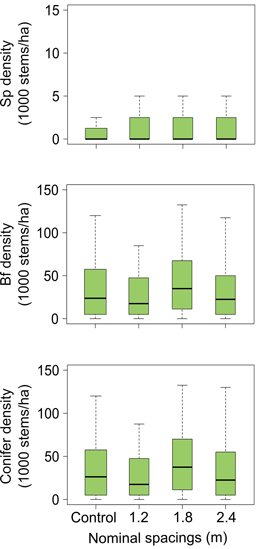
Fig. 5. Estimated values (lower hinge = within 1.5 times interquartile range below first quartile, lower end of box = first quartile, middle of box = median, upper end of box = third quartile, upper hinge = within 1.5 times interquartile range above third quartile) for stem density in next rotation balsam fir stands as influenced by different intensities of precommercial thinning in previous stands. Note the differences in y-axes range. Sp = spruce; Bf = balsam fir.
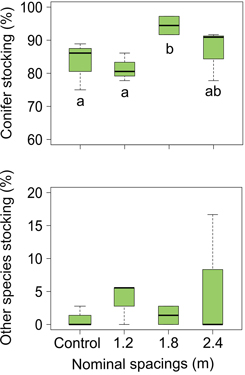
Fig. 6. Estimated values (lower hinge = within 1.5 times interquartile range below first quartile, lower end of box = first quartile, middle of box = median, upper end of box = third quartile, upper hinge = within 1.5 times interquartile range above third quartile) for stocking in next rotation balsam fir stands as influenced by different intensities of precommercial thinning in previous stands. Conifer stocking boxplot shows result of multiple comparison for significance. Note the differences in y-axes range.
4 Discussion
We predicted that silviculture scenarios with PCT would have legacy effects on the density and growth of regenerating conifers of next rotation stands; density, stocking and growth were expected to be positively correlated with increasing PCT intensity. Our results did not entirely support our hypothesis: PCT did not significantly influence regeneration size or density of the next rotation stands, but it significantly affected conifer stocking (Table 1). Indeed, conifer stocking from thinned stands using 1.8 m nominal spacing was higher than conifer stocking from both control stands and thinned stands resulting from the 1.2 m nominal spacing (Fig. 6), a result in line with the optimal PCT intensity in term of individual tree and stand growth (1.8 m nominal spacing; Pitt et al. 2013b). However, the significant higher conifer stocking found in the 1.8 m spacing treated stands does not have any implication for silviculture or forest management, as both treated and untreated stands were fully stocked (conifer stocking median > 80%; Fig. 6) (Pominville and Ruel 1995).
We interpret the lack of legacy effects of PCT on next rotation stand regeneration as being related, at least in part, to the shade tolerance of balsam fir (Humbert et al. 2007). Given that the species can establish an abundant seedling bank in the understory of closed canopy stands (Frank 1990; Parent and Ruel 2002), it appears that the initial differences in canopy closure that existed between the thinned and unthinned plots (until about 30 y of age; Pitt and Lanteigne 2008) did not influence regeneration density. Also, about 43 y after PCT was applied, the treated and control stands had converged to similar densities and live crown ratios due to self-thinning (Pitt and Lanteigne 2008), hence limiting potential differences in terms of available seed trees (Frank 1990; Ontario Ministry of Natural Resources 2000) and understory vegetation at maturity (Hale 2003; Cole et al. 2008, 2010). The lack of treatment effect on regeneration density and on competing vegetation cover probably explains why seedling growth (as expressed by tree size) did not vary significantly; intraspecific competition was similar among treatments and interspecific competition by broadleaf species did not vary across treatments as well (Table 1). The cover of competing vegetation was low (<10%), as was other species (mostly hardwood) density and stocking (median = 0stems ha–1; <6%). Indeed, all the tested scenarios included the application of glyphosate three years after harvesting, which was the year before we conducted the vegetation and regeneration survey. Glyphosate is a broad-spectrum post‐emergence non‐selective herbicide (Grossbard and Atkinson 1985) known for its efficacy at controlling hardwood species and favoring conifer dominance when applied in late summer/early fall (Pitt et al. 2004). Our results thus confirm that the vegetation management strategy used on these sites was effective at limiting interspecific competition and favoring balsam fir.
The abundance of CWD, which can impact regeneration and competing vegetation development (Trottier-Picard et al. 2014), did not vary significantly as a function of PCT intensity; overall slash and large woody debris cover were equivalent among treatments (Table 1). Although initial stocking levels should influence the accumulation and overall abundance of CWD (Sturtevant et al. 1997), it appears that the nominal spacings used during PCT were not different enough to significantly affect the abundance of CWD of the managed stands. This result is also in line with the convergence of mature stand density due to self-thinning (Pitt and Lanteigne 2008).
4.1 Management implications
Our study suggests that precommercial thinning of dense balsam fir stands has few, if any legacy effects on next rotation stand regeneration when applied within the range in intensity and timing we have tested. This silviculture practice, known to enhance diameter growth and volume of individual stems, enhance harvested piece size and products value, control stand composition and reduce rotation length and harvesting costs (Pothier 2002; Pitt and Lanteigne 2008; Zhang et al. 2009; Prévost and Gauthier 2012; Pitt et al. 2013b), should not impair next rotation stand regeneration.
However, balsam fir is a cold-adapted boreal species that is at high-risk under the predicted future climate conditions, particularly in the Acadian region (Bourque and Hassan 2008; Taylor et al. 2017). Hence, future vegetation management and tending treatments (e.g. release, PCT) should preferentially increase tree diversity by selecting for healthy individuals of other species, including hardwoods, to support the resilience of these ecosystems to climate change (Millar et al. 2007; Nagel et al. 2017). Indeed, the tested scenarios have resulted in a clear dominance of balsam fir in the regeneration layer (median ~20 000 stems ha–1) and the near absence of other species (median ~0 stems ha–1), which raises concerns for the resilience of these stands in the long term.
Acknowledgements
We are indebted to the previous researchers, technicians, managers and institutions, private or governmental, who have contributed in establishing, maintaining and documenting the Green River study over the decades, including Dr. Gordon Baskerville, Dr. Doug Pitt and Len Lanteigne, as well as Acadian Timber Corp., Twin Rivers Paper Co. Inc., J.D. Irving Limited, the New Brunswick Department of Energy and Resource Development (NBDERD), and the Canadian Forest Service. We also thank Martin Noel (NBDERD) for insightful discussions, and Shawn Palmer, Jim Coleman and Glen Forbes for their expertise and careful field measurements.
References
Baldwin V.C. (1977). Regeneration following shelterwood cutting in a New Brunswick softwood stand. In: Information Report M-X-76. Maritime Forest Research Center, Canadian Forest Service, Fredericton, New Brunswick. 23 p.
Baskerville G.L., Hughes E.L., Loucks O.L. (1960). Research by the Federal Forestry Branch in the Green River Project. The Forestry Chronicle 36(3): 265–277. https://doi.org/10.5558/tfc36265-3.
Bicho P., Portillo E., Yuen B., Yan D., Pitt D.G. (2013). Effects of precommercial thinning on the forest value chain in northwestern New Brunswick: part 5 – kraft and thermomechanical pulping and pulp quality. The Forestry Chronicle 89(04): 490–501. https://doi.org/10.5558/tfc2013-090.
Bourque C.P.-A., Hassan Q.K. (2008). Projected impacts of climate change on species distribution in the Acadian Forest region of eastern Nova Scotia. The Forestry Chronicle 84(4): 553–557. https://doi.org/10.5558/tfc84553-4.
Brissette J.C. (1996). Effects of intensity and frequency of harvesting on abundance, stocking and composition of natural regeneration in the Acadian forest of eastern North America. Silva Fennica 30(2–3): 301–314. https://doi.org/10.14214/sf.a9242.
Canadian Council of Forest Ministers (2019). http://nfdp.ccfm.org/en/data/regeneration.php. [Cited 28 April 2019]
Cole E.C., Hanley T.A., Newton M. (2010). Influence of precommercial thinning on understory vegetation of young-growth Sitka spruce forests in southeastern Alaska. Canadian Journal of Forest Research 40(4): 619–628. https://doi.org/10.1139/X10-009.
Cole H.A., Newmaster S.G., Lanteigne L., Pitt D. (2008). Long-term outcome of precommercial thinning on floristic diversity in north western New Brunswick, Canada. iForest 1: 145–156. https://doi.org/10.3832/ifor0470-0010145.
Côté S. (2013). Le sapin baumier [Balsam fir]. In: Le guide sylvicole du Québec : Les fondements biologiques de la sylviculture. [Silvicultural Guide of Quebec: Biological Foundations of Silviculture]. Volume 1. Les Publications du Québec, Québec. p. 120–123.
Côté S., Bélanger L. (1991). Variations de la régénération préétablie dans les sapinières boréales en fonction de leurs caractéristiques écologiques. [Variations in advanced regeneration in boreal balsam fir stands based on their ecological characteristics]. Canadian Journal of Forest Research 21(12): 1779–1795. https://doi.org/10.1139/x91-246.
Department of Natural Resources of New Brunswick (2007). Our landscape heritage: the story of ecological land classification in New Brunswick. Second Edition. Fredericton, New Brunswick. 359 p.
Department of Natural Resources of New Brunswick (2008). Silviculture statistics crown lands. No 2008–02. 14 p. https://www2.gnb.ca/content/dam/gnb/Departments/nr-rn/pdf/en/ForestsCrownLands/CrownSivilStats.pdf.
Duchesne I., Pitt D.G., Tanguay F. (2013). Effects of precommercial thinning on the forest value chain in northwestern New Brunswick: part 4 – lumber production, quality and value. The Forestry Chronicle 89(04): 474–489. https://doi.org/10.5558/tfc2013-089.
Environment Canada (2010). Canadian Climate Normals 1981–2010 station data. http://climate.weather.gc.ca/climate_normals/index_e.html. [Cited 16 Nov 2018].
Forest Management Branch (2004). Forest management manual for New Brunswick crown land. Natural Resources, Fredericton, New Brunswick. 137 p.
Frank R.M. (1990). Abies balsamea (L.) Mill. Balsam Fir. In: Silvics of North America: 1. conifers. Agriculture Handbook 654, United States Department of Agriculture, Forest Service, Washington, DC. p. 26–35.
Fraver S., Wagner R.G., Day M. (2002). Dynamics of coarse woody debris following gap harvesting in the Acadian forest of central Maine, USA. Canadian Journal of Forest Research 32(12): 2094–2105. https://doi.org/10.1139/x02-131.
Gauthier M.-M., Tremblay S. (2018). Precommercial thinning as a silvicultural option for treating very dense conifer stands. Scandinavian Journal of Forest Research 33(5): 446–454. https://doi.org/10.1080/02827581.2017.1418422.
Gauthier S., Bernier P., Burton P.J., Edwards J., Isaac K., Isabel N., Jayen K., Le Goff H., Nelson E.A. (2014). Climate change vulnerability and adaptation in the managed Canadian boreal forest. Environmental Reviews 22(3): 256–285. https://doi.org/10.1139/er-2013-0064.
Grossbard E., Atkinson D. (1985). The Herbicide Glyphosate. Butterworths and Compagny Ltd, London. 490 p. ISBN 0-408-11153-4.
Hale S.E. (2003). The effect of thinning intensity on the below-canopy light environment in a Sitka spruce plantation. Forest Ecology and Management 179(1–3): 341–349. https://doi.org/10.1016/S0378-1127(02)00540-6.
Hothorn T., Bretz F., Westfall P., Heiberger R.M., Schuetzenmeister A., Scheibe S. (2019). Package “multcomp”. https://cran.r-project.org/web/packages/multcomp/multcomp.pdf. [Cited 28 April 2019].
Humbert L., Gagnon D., Kneeshaw D., Messier C. (2007). A shade tolerance index for common understory species of northeastern North America. Ecological Indicators 7(1): 195–207. https://doi.org/10.1016/j.ecolind.2005.12.002.
Lindgren P.M.F., Ransome D.B., Sullivan D.S., Sullivan T.P. (2006). Plant community attributes 12 to 14 years following precommercial thinning in a young lodgepole pine forest. Canadian Journal of Forest Research 36(1): 48–61. https://doi.org/10.1139/x05-228.
Loucks O.L. (1962). A forest classification for the Maritime provinces. The Proceedings of the Nova Scotian Institute of Science 25: 86–167.
Millar C.I., Stephenson N.L., Stephens S.L. (2007). Climate change and forests of the future: managing in the face of uncertainty. Ecological Applications 17(8): 2145–2151. https://doi.org/10.1890/06-1715.1.
Nagel L.M., Palik B.J., Battaglia M.A., D’Amato A.W., Guldin J.M., Swanston C.W., Janowiak M.K., Powers M.P., Joyce L.A., Millar C.I., Peterson D.L., Ganio L.M., Kirschbaum C., Roske M.R. (2017). Adaptive silviculture for climate change: a national experiment in manager-scientist partnerships to apply an adaptation framework. Journal of Forestry 115(3): 167–178. https://doi.org/10.5849/jof.16-039.
Nyland R.D., Kenefic L.S., Bohn K.K., Stout S.L. (2016). Silviculture: concepts and applications. Third edition. Waveland Press Inc., Long Grove, Illinois. 680 p. ISBN 1-4786-2714-X.
Ontario Ministry of Natural Resources (2000). Appendix B: autecology of southern Ontario tree species. In: A silvicultural guide to managing southern Ontario forests. Technical series. p. 451–529.
Parent S., Ruel J.-C. (2002). Chronologie de la croissance chez des semis de sapin baumier (Abies balsamea (L.) Mill.) après une coupe à blanc avec protection de la régénération. [Chronology of balsam fir (Abies balsamea (L.) Mill.) seedlings growth after clearcut harvesting with protection of advanced regeneration]. The Forestry Chronicle 78(6): 876–885. https://doi.org/10.5558/tfc78876-6.
Piene H. (1978). Effects of increased spacing on carbon mineralization rates and temperature in a stand of young balsam fir. Canadian Journal of Forest Research 8(4): 398–406. https://doi.org/10.1139/x78-059.
Pinheiro J., Bates D., R-core (2018). Package “nlme”. https://cran.r-project.org/web/packages/nlme/nlme.pdf. [Cited 28 April 2019].
Pitt D., Lanteigne L. (2008). Long-term outcome of precommercial thinning in northwestern New Brunswick: growth and yield of balsam fir and red spruce. Canadian Journal of Forest Research 38(3): 592–610. https://doi.org/10.1139/X07-132.
Pitt D.G., Wagner R.G., Towill W.D. (2004). Ten years of vegetation succession following ground-applied release treatments in young black spruce plantations. Northern Journal of Applied Forestry 21(3): 123–134. https://doi.org/10.1093/njaf/21.3.123.
Pitt D.G., Lanteigne L., Hoepting M.K., Farrell J. (2013a). Effects of precommercial thinning on the forest value chain in northwestern New Brunswick: a fifty-year legacy of forest research continues. The Forestry Chronicle 89(4): 439–445. https://doi.org/10.5558/tfc2013-085.
Pitt D.G., Lanteigne L., Hoepting M.K., Plamondon J. (2013b). Effects of precommercial thinning on the forest value chain in northwestern New Brunswick: part 1 – roundwood production and stumpage value. The Forestry Chronicle 89(04): 446–457. https://doi.org/10.5558/tfc2013-086.
Pitt D., Lanteigne L., Hoepting M.K., Plamondon J., Duchesne I., Bicho P., Warren G. (2013c). Effects of precommercial thinning on the forest value chain in northwestern New Brunswick: part 6 – estimating the economic benefits. The Forestry Chronicle 89(04): 502–511. https://doi.org/10.5558/tfc2013-091.
Plamondon J., Pitt D.G. (2013). Effects of precommercial thinning on the forest value chain in northwestern New Brunswick: part 2 – efficiency gains in cut-to-length harvesting. The Forestry Chronicle 89(04): 458–463. https://doi.org/10.5558/tfc2013-087.
Pominville P., Ruel J.-C. (1995). Effets de la coupe à blanc et de la coupe par bandes sur la régénération obtenue après 5 ans dans des pessières noires du Québec. [Effects of clearcut harvesting and strip cutting on regeneration 5 years post-harvesting in black spruce stands in Quebec]. Canadian Journal of Forest Research 25(2): 329–342. https://doi.org/10.1139/x95-037.
Pothier D. (2002). Twenty-year results of precommercial thinning in a balsam fir stand. Forest Ecology and Management 168(1): 177–186. https://doi.org/10.1016/S0378-1127(01)00738-1.
Pothier D., Margolis A. (1991). Analysis of growth and light interception of balsam fir and white birch saplings following precommercial thinning. Annals of Forest Science 48(2): 123–132. https://doi.org/10.1051/forest:19910201.
Prévost M., Gauthier M.-M. (2012). Precommercial thinning increases growth of overstory aspen and understory balsam fir in a boreal mixedwood stand. Forest Ecology and Management 278: 17–26. https://doi.org/10.10163j.foreco.2012.05.005.
R Core Team (2018). R: a language and environment for statistical computing. http://www.R-project.org/. [Cited 28 April 2019]
Rowe J.S. (1972). Forest regions of Canada. Department of Environment, Canadian Forest Service. Information Canada Fo47-1300F. 172 p.
Ruel J.-C. (1989). Importance de la régénération préexistante dans les forêts publiques du Québec. [Importance of advanced regeneration on Quebec public lands]. Annals of Forest Science 46(4): 345–359. https://doi.org/10.1051/forest:19890403.
Ruel J.-C., Messier C., Claveau Y., Doucet R., Comeau P. (2000). Morphological indicators of growth response of coniferous advance regeneration to overstorey removal in the boreal forest. The Forestry Chronicle 76(4): 633–642. https://doi.org/10.5558/tfc76633-4.
Steel R.G.D., Torrie J.H., Dickey D.A. (1997). Principles and procedures of statistics: a biometrical approach. Third edition. McGraw-Hill Inc, New York. 666 p. ISBN 0-07-061028-2.
Sturtevant B.R., Bissonette J.A., Long J.N., Roberts D.W. (1997). Coarse woody debris as a function of age, stand structure, and disturbance in boreal Newfoundland. Ecological Applications 7(2): 702–712. https://doi.org/10.1890/1051-0761(1997)007[0702:CWDAAF]2.0.CO;2.
Taylor A.R., Boulanger Y., Price D.T., Cyr D., McGarrigle E., Rammer W., Kershaw Jr. J.A. (2017). Rapid 21st century climate change projected to shift composition and growth of Canada’s Acadian Forest Region. Forest Ecology and Management 405: 284–294. https://doi.org/10.1016/j.foreco.2017.07.033.
Thibodeau L., Raymond P., Camiré C., Munson A.D. (2000). Impact of precommercial thinning in balsam fir stands on soil nitrogen dynamics, microbial biomass, decomposition, and foliar nutrition. Canadian Journal of Forest Research 30(2): 229–238. https://doi.org/10.1139/x99-202.
Trottier-Picard A., Thiffault E., DesRochers A., Paré D., Thiffault N., Messier C. (2014). Amounts of logging residues affect planting microsites: a manipulative study across northern forest ecosystems. Forest Ecology and Management 312: 203–215. https://doi.org/10.1016/j.foreco.2013.10.004.
Zeide B. (2001). Thinning and growth: a full turnaround. Journal of Forestry 99(1): 20–25. https://doi.org/10.1093/jof/99.1.20.
Zhang S., Chauret G., Tong Q. (2009). Impact of precommercial thinning on tree growth, lumber recovery and lumber quality in Abies balsamea. Scandinavian Journal of Forest Research 24(5): 425–433. https://doi.org/10.1080/02827580903124392.
Total of 53 references.
Send to email
