Amy Wotherspoon  ,
Nelson Thiffault,
Robert L. Bradley
,
Nelson Thiffault,
Robert L. Bradley
Resource availability and physiological response of black spruce to scarification in two climatic regions of Québec (Canada)
Wotherspoon A., Thiffault N., Bradley R. L. (2020). Resource availability and physiological response of black spruce to scarification in two climatic regions of Québec (Canada). Silva Fennica vol. 54 no. 4 article id 10375. https://doi.org/10.14214/sf.10375
Highlights
- Mechanical scarification of clearcut sites in two climatic regions maintained high growth rates of regenerating black spruce 18 years after treatment
- In both regions, scarification reduced soil moisture and increased water use efficiency
- In the cooler-humid region, scarification improved soil nitrogen mineralization.
Abstract
Scarification is a mechanical site preparation technique designed to create microsites that will favor the growth of planted tree seedlings after clearcutting. However, the positive growth response of black spruce (Picea mariana (Mill.) Britton, Sterns & Poggenb.) to scarification varies across different sites. We hypothesized that this was due to different forms of physiological stress induced by different climates or by the severity of competition from ericaceous shrubs. We thus compared the effects of scarification on black spruce needle gas exchange and other foliar properties, as well as on indices of soil water and nitrogen availability, in relatively warm-dry (Abitibi) vs. cool-humid (Côte-Nord) climates in the province of Québec (Canada). We found a similar positive effect of scarification on tree height in Abitibi and Côte-Nord. Scarification reduced soil moisture in both climatic regions, but increased soil N mineralization in Côte-Nord only. Accordingly, scarification increased the instantaneous water use efficiency in both climate regions, but decreased photosynthetic N use efficiency in Côte-Nord only. In both regions, we found a positive relationship between foliar δ18O and δ13C on scarified plots, providing further evidence that increased growth due to scarification depends on a decrease in stomatal conductance. We conclude that scarification increases total evapotranspiration of trees evenly across the east-to-west moisture gradient in the province of Québec, but also improves long-term soil nutritional quality in a cooler-humid climate.
Keywords
Picea mariana;
foliar δ18O;
foliar δ13C;
needle gas exchange;
photosynthetic nitrogen use efficiency;
water use efficiency
-
Wotherspoon,
Département de biologie, Université de Sherbrooke, Sherbrooke, Québec, J1K 2R1, Canada
E-mail
amy.wotherspoon@usherbrooke.ca
- Thiffault, Canadian Wood Fibre Centre, Canadian Forest Service, Natural Resources Canada, 1055 du. P.E.P.S., P.O. Box 10380, Québec, G1V 4C7, Canada E-mail nelson.thiffault@canada.ca
- Bradley, Département de biologie, Université de Sherbrooke, Sherbrooke, Québec, J1K 2R1, Canada E-mail robert.bradley@usherbrooke.ca
Received 13 May 2020 Accepted 12 August 2020 Published 26 August 2020
Views 61790
Available at https://doi.org/10.14214/sf.10375 | Download PDF

1 Introduction
Scarification is a mechanical site preparation technique used by foresters after clearcutting in order to create microsites that will favor the growth of planted tree seedlings. Scarifiers consist of motorized disks with multiple blades that rip and lift surface organic layers, thus exposing the mineral soil along trenches in which tree seedlings are planted. Positive effects of scarification on tree seedling growth have been ascribed to decreased competition from vegetation (Hébert et al. 2014), increased soil temperature (Thiffault and Jobidon 2006; Sikström et al. 2020), improved light availability for seedlings (Willis et al. 2015), improved soil drainage (Rappe George et al. 2017) and increased soil nutrient mineralization (Johansson 1994). In the province of Québec (Canada), scarification is commonly used to improve the regeneration of planted black spruce (Picea mariana (Mill.) Britton, Sterns & Poggenb.) stands, the province’s most economically important boreal tree species. While the growth response of black spruce to scarification is generally positive, its effectiveness varies across sites. For example, Hébert et al. (2014) found that scarification had a greater positive effect on seedling growth in black spruce–feather moss forest sites that had been clearcut compared to patches of unmanaged open lichen woodland. Understanding how such site-specific characteristics affect the physiology of black spruce could help us predict the relative growth response to mechanical scarification in different regions.
In Québec, black spruce has a wide geographic distribution spanning several bioclimatic zones, resulting in different stand structures and understory plant communities (Nicault et al. 2015). For example, the Abitibi region located in western Québec is characterized by a relatively warm-dry summer climate compared to the Côte-Nord region located on the northeast coast of the St-Lawrence River. Regenerating black spruce cutovers in the Abitibi region benefit from more degree-days during the growing season and appear to have less competing vegetation than those in the Côte-Nord region. That is because the relatively cool-humid climate of the Côte-Nord region often results in a dense cover of ericaceous shrubs such as Kalmia angustifolia L. and Rhododendron groenlandicum (Oeder) Kron & Judd. While these shrub species are also found in the Abitibi region, they are more prone to dominate on clearcuts in the Côte-Nord region resulting in the growth check of regenerating black spruce seedlings (Reicis et al. 2020). Thus, releasing black spruce seedlings from the competing vegetation through mechanical scarification may be more beneficial in Côte-Nord than in Abitibi.
We posit that scarification in Abitibi and Côte-Nord exacerbates or alleviates different forms of physiological stress induced by climate or by competing vegetation. For example, soil water is a common limiting resource for many boreal tree species, including black spruce (Walker et al. 2015). In the relatively warm-dry Abitibi region, faster growth rates on scarified plots should increase total evapotranspiration of the regenerating stand and increase the needle-air vapor pressure deficit (VPD), thereby depleting soil moisture. If an increase in water-stress occurs on scarified plots, then black spruce would need to reduce stomatal conductance to limit water loss and increase water use efficiency (WUE) in order to maintain growth rates (Flanagan and Farquhar 2014). We should observe, therefore, an increase in WUE following scarification in the Abitibi region. Such moisture deficits on scarified plots are less likely to occur in the Côte-Nord region because of higher annual precipitation and lower potential evapotranspiration.
Another way by which scarification could have different effects on black spruce physiology in Abitibi and Côte-Nord is by having different effects on soil fertility. The dense ericaceous shrub cover that characterizes cool-moist climates (e.g. Côte-Nord) can result in low soil N fertility (Bradley et al. 1997). This is generally ascribed to the high concentrations of condensed tannins in ericaceous leaf litter, which prevents soil N from mineralizing (Joanisse et al. 2009). In such N-limited environments, black spruce seedlings need to increase their rates of photosynthesis per unit of foliar N (i.e. photosynthetic N use efficiency (PNUE)) in order to maintain growth rates. If scarification increases tree growth in the Côte-Nord region, then we expect that this will improve soil N fertility in two ways: (1) by reducing ericaceous tannin concentrations due to shading (Hofland-Zijlstra and Berendse 2009); (2) by adding proportionately more conifer needle litter relative to ericaceous litter to the forest floor (Joanisse et al. 2007). We should observe, therefore, a decrease in PNUE following scarification in the Côte-Nord region.
We report on a study where we measured needle gas exchange and other foliar properties, as well as indices of soil water and N availability, in regenerating black spruce plots in the Abitibi and Côte-Nord regions. The plots had been clearcut and scarified approximately 18 years prior to our study. At four dates throughout the growing season, we calculated instantaneous water use efficiency (iWUE) and PNUE using leaf level gas exchange measurements. At the end of the growing season, we used leaf oxygen and carbon isotopic ratios to calculate seasonal water use efficiency (sWUE). Based on the rationale presented above, we predicted that: (1) scarification would have a relatively larger positive effect on black spruce growth in Côte-Nord compared to Abitibi; (2) scarification in Abitibi would increase moisture stress and WUE relative to non-scarified plots; (3) scarification in Côte-Nord would increase soil N fertility and reduce PNUE relative to non-scarified plots.
2 Methods
2.1 Study sites
The two experimental sites are located 573 km apart, at opposite limits of a west-to-east gradient of increasing precipitation and decreasing summer temperature in the province of Québec. The Abitibi site (48°30´N, 76°56´W, altitude of 396 m a.s.l.) is located in the balsam fir (Abies balsamea (L.) Mill.) – paper birch (Betula papyrifera Marshall) bioclimatic domain (Saucier et al. 2009) and is characterized by a 30-year mean annual temperature of 2.5 °C and an annual precipitation range of 800–900 mm, 30% falling as snow (Blouin and Berger 2002). The region has 1443 degree (°C) days per year and a 30-year mean temperature of 16 °C during the three warmest months of the year (June to August). The growing season in Abitibi lasts approximately 150–160 days (Environment Canada 2018). The Côte-Nord site (49°47´N, 69°17´W, altitude of 472 m a.s.l.) is located in the black spruce–feather moss bioclimatic domain and is characterized by a mean annual temperature of –1.0 °C and an annual precipitation range of 1100–1300 mm, with 34–40% falling as snow (Morneau and Landry 2007). The region has 1179 degree (°C) days per year and a 30-year mean temperature of 14.2 °C during the three warmest months of the year. The growing season in Côte-Nord lasts approximately 140 days (Environment Canada 2018).
Soils on both sites are classified as haplohumod spodosols (Soil Survey Staff 2006) developed on well-drained glacial till, with clay contents ≤ 4%. Mean forest floor thickness is 8 cm in Abitibi and 18 cm in Côte-Nord. In Abitibi, the understory shrub layer is relatively sparse and includes K. angustifolia and Vaccinium spp., with Cladina spp. lichen as the ground cover. In Côte-Nord, the understory shrub layer is relatively dense and consists mainly of K. angustifolia and R. groenlandicum, with feather moss (Pleurozium schreberi (Willd. ex Brid.) Mitt.) as the ground cover.
The Abitibi and Côte-Nord sites had been clearcut harvested in 1999 and 1993 respectively. On each site, single-pass scarified and non-scarified treatment plots were established in 1999 using a TTS disk trencher. Plots in Abitibi measured 15 m × 10 m, whereas those in Côte-Nord measured 32 m × 6.5 m. At each site, scarified trenches (ca. 60 cm wide, 30 cm deep) were established lengthwise at 2 m apart, resulting in five rows per plot in Abitibi and three rows per plot in Côte-Nord. On each site, both treatments were replicated in five blocks within a 2.5 ha area in Abitibi and a 5.0 ha area in Côte-Nord. Treatment plots within each block were separated by a 6 m buffer strip in Abitibi and by a 2 m buffer strip in Côte-Nord. In June 2000, each plot in Abitibi and Côte-Nord were respectively planted with 18 and 32 two-year-old black spruce seedlings, grown from seed in 110 mL containers, with a 2.5 m distance between seedlings (ca. 1200 and 1500 seedlings ha–1 respectively). Seedlings planted in Abitibi were grown at a nursery in the town of Trécesson, whereas those planted in Côte-Nord were grown at a nursery in the town of Forestville (Thiffault et al. 2005; Thiffault and Jobidon 2006).
2.2 Black spruce growth
For three consecutive years (2015 to 2017), the height of black spruce trees in each plot was measured at the end of each growing season (mid-September) using a telescopic meter stick. From these values, we calculated a two-year relative growth rate (RGR) for each tree as:
![]()
2.3 Forest floor sampling and analyses
At the start of the 2017 sampling season, diagonal transects were established on each plot at each site. Twelve sampling points per plot were established along three transects in Abitibi and along four transects in Côte-Nord. On four monthly sampling dates between May and August 2017, forest floor subsamples (ca. 0.5 l) were collected at each sampling point in each plot. Forest floor subsamples were bulked into three composite samples per plot, sieved through a 5 mm metal mesh screen, placed into 3.8 l plastic bags and transported in coolers to the laboratory (Université de Sherbrooke). Forest floor samples were stored at 4 °C until analyzed.
Gravimetric moisture content of each forest floor sample was calculated based on the weight loss of subsamples after drying for one week at 60 °C in an air-draft oven. The subsamples were then ground with a ball mill and analyzed for total N by high temperature combustion and thermo-conductometric detection, using a Vario Macro dry combustion analyzer (Elementar Analysensysteme GmbH, Hanau, Germany).
Fresh forest floor subsamples (5 g dry mass equiv.) were placed in covered jars and incubated for 30 days under aerobic conditions. Following the incubation, the forest floor subsamples were extracted in aqueous 1 N KCl and analyzed colorimetrically for NH4+-N concentration (salicylate-nitroprusside-hypochlorite assay) using a continuous flow analyzer (Astoria2, Astoria-Pacific International, Clackamas, OR). Given that scarification results in a mixing of soil horizons, forest floor samples can sometimes be contaminated with small amounts of mineral soil, which has a higher specific density than forest floor material. For this reason, mineralizable N was expressed relative to total N rather than to sample dry weight (see Discussion).
2.4 Gas exchange measurements
At each forest floor sampling date in 2017, gas exchange measurements were performed on 1-year-old needles from the upper-third of the canopy of two trees in each plot. Measurements were performed between 10:00 a.m. and 2:00 p.m. local time using a LiCor 6400XT Portable Photosynthesis System (LiCor Scientific, Lincoln, NE). Selected twigs were inserted into a conifer needle chamber with saturating light (1000 µmol photons m–2 s–1) provided by a mixed red/blue LED. The CO2 concentration in the chamber was maintained at 400 ppm with a flow rate of 150 µmol s–1. Needles were left in the chamber for 5 min prior to measurement to allow gas exchange rates to equilibrate. The measured variables included net rates of CO2 assimilation (i.e. photosynthesis) and water vapor loss (i.e. transpiration (E)). Following each measurement, twigs were excised, sealed into small plastic bags, and returned on ice to the laboratory. Needles were scanned and their projected area was calculated using WinSEEDLE image analysis software (Regent Instruments, Québec City, QC). Needles were then dried, weighed and analyzed for total N, as described above. From these values, we calculated net rate of photosynthesis per needle surface area (Pn) and per needle mass (A). Instantaneous water use efficiency was calculated as:
![]()
Photosynthetic N use efficiency was calculated as:
![]()
2.5 Needle sampling and analyses
Unlike instantaneous leaf-level gas exchange measurements made on only a few sampling dates, foliar δ13C and δ18O isotope ratios reflect season-long gas exchange fluxes (Farquhar and Lloyd 1993). An increase in δ13C reflects an increase in the 13C/12C ratio of plant biomass, which may occur for one of two reasons: (1) lower stomatal conductance (i.e. higher WUE) resulting in a decrease of intercellular CO2, allowing more of the heavier 13C-CO2 to diffuse to chloroplasts and be fixed into plant biomass; (2) higher net rates of photosynthesis at a given stomatal conductance, resulting in a faster diffusion of CO2 to chloroplasts and a decrease of intercellular CO2. To determine which of these two mechanisms is most prevalent, we must look at variations in foliar δ18O. An increase in the total evapotranspiration of a given tree will result in a preferential loss of the lighter 16O-H2O and an increase in the assimilation of the heavier 18O isotope into plant biomass. Hence, a significant positive relationship between foliar δ13C and δ18O isotope values indicates that variations in δ13C values are likely induced through moisture stress and controlled by stomatal conductance rather than by photosynthetic capacity.
In September 2016, needle samples were collected from two trees from each plot in order to quantify δ13C and δ18O isotope ratios. We pooled an equal amount of ground needle material from both trees in each plot, and sent a 20 g subsample to the G.G. Hatch Stable Isotopes Laboratory at the University of Ottawa (Canada). The 20 subsamples were flash combusted at 1450 °C (Dumas combustion) and the resulting gas products were analyzed for their δ18O and δ13C isotope ratios using a Delta V Advantage isotope ratio mass spectrometer (Thermo Scientific, Bremen, Germany).
At each forest floor sampling date in 2017, needles were collected from the same two black spruce trees in each plot, in all orientations from the top third of the canopy, using pruning shears. Needle samples were placed in coolers under ice packs and transported to the laboratory, where they were oven-dried at 60 °C. Dried needle samples were ground in a ball mill and analyzed for total N, as described above.
2.6 Statistical Analyses
The effects of scarification and sites, as well as their interactions, on the various response variables were tested using mixed effects models. For tree height and RGR, the identity of each block was used as a random variable. For soil properties, the identity of each block and the sampling date were used as random variables. For gas exchange and foliar measurements, the identity of each block, the sampling date and the identity of each tree were used as random variables. When significant interactions were found between scarification and site, the effects of scarification were tested within each site. Linear regression was used to test the relationship between δ18O and δ13C isotopic values. All statistical analyses were performed using the lmer package in R statistical software (Version 3.2.2, R Development Core Team 2015). A significance level of α = 0.05 was used, unless otherwise stated.
3 Results
3.1 Tree height and relative growth rate
We found a significant (P < 0.001) effect of scarification on tree height (Fig. 1a), with average tree height (n = 250) being 136% greater on scarified than non-scarified plots across both sites. Likewise, from 2015 to 2017, RGR across both sites was 39% greater (P < 0.001) in scarified than non-scarified plots (Fig. 1b).
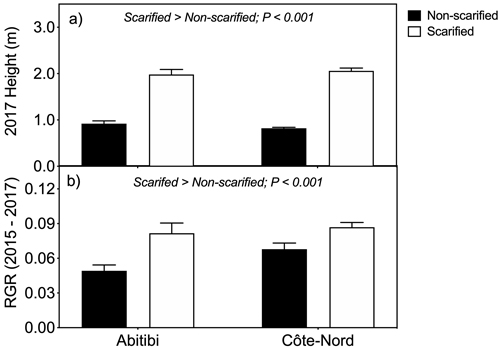
Fig. 1. Tree height (a) and relative growth rate (RGR) (b) of black spruce trees on non-scarified and scarified plots, at two boreal sites in Québec. Trees were planted as containerized 2-year-old seedlings in 1999. Two way mixed model ANOVA revealed a significant (P < 0.001) effect of scarification on tree height and RGR across both sites (N = 250 trees per treatment across sites). Vertical lines denote standard errors of the means (n = 90 in Abitibi; n = 160 in Côte-Nord).
3.2 Forest floor moisture and N mineralization
On both sites, scarification significantly (P < 0.001) reduced forest floor gravimetric moisture content across sampling dates (Fig. 2a). We found a significant (P = 0.013) site × scarification interaction on forest floor N mineralization across sampling dates. More specifically, N mineralization (per total N) in Côte-Nord was significantly higher (P = 0.037) in scarified than non-scarified plots, whereas no effect was observed in Abitibi (Fig. 2b).
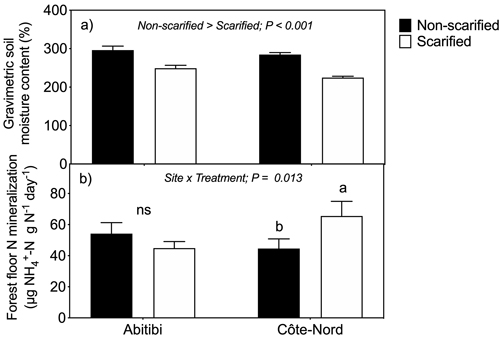
Fig. 2. Forest floor gravimetric moisture content (a) and nitrogen mineralization rate per total N (b) in non-scarified and scarified plots at two boreal sites in Québec. Each bar represents mean values based on 3 bulk samples × 5 plots × 4 sampling dates (n = 60). Two way mixed model ANOVA revealed a significant (P < 0.001) main effect of treatment on forest floor moisture content (Fig. 2a). In the case of N mineralization (Fig. 2b), a significant site × treatment interaction required us test the effects of scarification within each site; consequently, statistically different (P = 0.037) means were found in Côte-Nord (different lower-case letters), but not in Abitibi (ns = not significant). Vertical lines denote standard errors of the means.
3.3 Foliar measurements
Net rate of photosynthesis per needle surface area (Pn) was higher in non-scarified than in scarified plots (Table 1), although this effect was not statistically significant (P = 0.087). Net rate of photosynthesis per needle mass (A) was significantly higher (P = 0.027) in Abitibi than in Côte-Nord. Transpiration (E) was significantly higher (P = 0.027) in non-scarified than in scarified plots. Scarification reduced foliar N in Abitibi and increased foliar N in Côte-Nord, although this interaction was not statistically significant (P = 0.087). The iWUE across both sites was significantly higher (P < 0.001) in scarified than in non-scarified plots (Fig. 3a), as well as in Abitibi compared to Côte-Nord (P < 0.001). Photosynthetic N use efficiency (PNUE) was significantly (P = 0.011) higher in Abitibi compared to Côte-Nord (Fig. 3b).
| Table 1. Mean gas exchange and foliar nitrogen measurements of one-year-old black spruce needles in scarified and non-scarified plots in two sites of Québec, 18 years after treatment. Trees were planted as containerized 2-year old seedlings in 1999. The reported treatment means are based on 2 trees × 5 plots × 4 sampling dates (n = 40). The reported P values result from two-way mixed model analyses of variance testing the effects of treatments and sites, as well as their interaction. Values in parentheses denote one standard error of the mean. | |||||
| Abitibi | Côte-Nord | Significant findings | |||
| Non-scarified | Scarified | Non-scarified | Scarified | ||
| Pn | 9.58 (0.90) | 8.92 (0.76) | 9.15 (0.98) | 6.75 (0.92) | Non-scarified > Scarified; (P = 0.087) |
| A | 0.033 (0.003) | 0.032 (0.003) | 0.025 (0.002) | 0.022 (0.002) | Abitibi > Côte-Nord; (P = 0.027) |
| E | 4.39 (0.61) | 3.00 (0.29) | 4.52 (0.43) | 3.84 (0.40) | Non-scarified > Scarified; (P < 0.003) |
| Foliar N | 7.13 (0.23) | 6.83 (0.22) | 6.90 (0.21) | 7.50 (0.18) | Site x Treatment; (P = 0.087) |
| Pn: CO2 exchange rate per needle area (µmol CO2 m2 s–1), A: CO2 exchange rate per needle mass (µmol CO2 g–1 s–1), E: transpiration (mmol H2O m–2 s–1), Foliar N: foliar nitrogen concentration (mg N g–1) | |||||
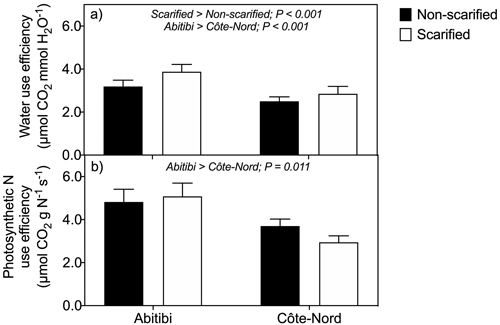
Fig. 3. Instantaneous water use efficiency (a) and photosynthetic nitrogen use efficiency (b) of black spruce trees on non-scarified and scarified plots, at two boreal sites in Québec. Trees were planted as containerized 2-year-old seedlings in 1999. Each bar represents mean values based on 2 trees × 5 plots × 4 sampling dates (n = 40). Two way mixed model ANOVA revealed significant main effects of treatments (P < 0.001) and sites (P < 0.001) on water use efficiency (Fig. 3a), and a significant effect of sites (P = 0.011) on photosynthetic nitrogen use efficiency (Fig. 3b).Vertical lines denote standard errors of the means.
Values of δ13C were greater in Côte-Nord than in Abitibi (P = 0.011). There was a significant (P = 0.012) site × treatment interaction on needle δ18O values (Fig. 4). More specifically, needle δ18O values were greater in Abitibi compared to Côte-Nord (P < 0.001), and significantly higher (P = 0.004) in scarified than non-scarified plots in the Abitibi site only. On both sites, there was a significant positive relationship between δ18O and δ13C on scarified plots (Abitibi: R2 = 0.51, P = 0.021; Côte-Nord: R2 = 0.49, P = 0.024), but not on the non-scarified plots (Fig. 4).
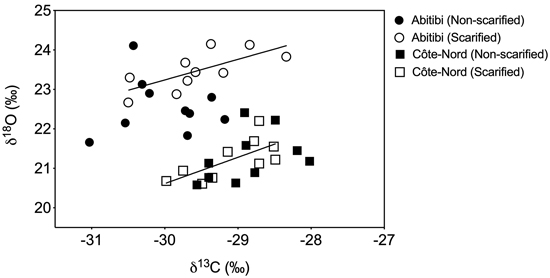
Fig. 4. Results of linear regression analysis testing the relationship between δ18O and δ13C isotopic values of one-year-old needles in non-scarified (closed symbols) and scarified (open symbols) plots, at two boreal sites in Québec. Needles were collected from two black spruce trees per plot, in September 2016 (n = 10). Trees were planted as containerized 2-year-old seedlings in 1999. We found a significant positive regression slope (solid line) on scarified plots only, on both the Abitibi (P = 0.021, R2 = 0.51) and the Côte-Nord (P = 0.024, R2 = 0.49) sites.
4 Discussion
Our data provided evidence that scarification increases black spruce height in both climatic regions of Québec. The fact that RGR was also higher on scarified plots suggests that scarification sustained favorable growth conditions, even 18 years after treatment. This implies a sustained release of black spruce trees from competing vegetation as well as a possible long-term increase in site nutritional quality. Contrary to what we had predicted, the relative increase in black spruce height was not higher in Côte-Nord than in Abitibi. We had based our prediction on the hypothesis that black spruce trees in Côte-Nord are subject to greater interference from ericaceous shrubs and would therefore benefit the most from scarification (as discussed below). What our data suggest, however, is that the relative growth response of black spruce to scarification may not necessarily relate only to the factors limiting growth before treatment, but also to the potential maximum growth rate of a given site. This potential growth rate is likely to be higher in Abitibi than in Côte-Nord, given the higher temperature and longer growing season. This is supported by models developed by Hamel et al. (2004), who found that the number of seasonal degree-days was the best predictor of black spruce productivity. Another factor that may boost the potential maximum growth rate in Abitibi is the fact that planting density was lower than in Côte-Nord (1200 vs. 1500 stems ha–1), resulting in lower intra-specific competition. Higher seasonal degree-days and/or lower intra-specific competition would both allow trees in Abitibi to benefit more from the favorable growth conditions created by scarification.
We initially hypothesized that faster growth rates on scarified plots would increase total stand evapotranspiration, and that this might lead to soil water deficits and moisture stress in the drier Abitibi climate, more so than in Côte-Nord. Likewise, we predicted that greater moisture stress on scarified plots would result in a greater increase in iWUE in Abitibi than in Côte-Nord. While we did observe a decrease in soil moisture and an increase of iWUE on scarified plots, the effect was similar on both sites. These data are consistent with the significantly lower transpiration rates in scarified plots on both sites (Table 1). Greater iWUE suggests that stomatal conductance may play an important role in mitigating moisture stress on scarified plots. This is corroborated by the significant positive relationships between foliar δ13C and δ18O on scarified plots at each site, but not on the non-scarified plots (Farquhar et al. 1998; Scheidegger et al. 2000). This implies that greater moisture stress caused by scarification at either site exceeded the difference in precipitation and soil water availability that occurred between the two sites.
As mentioned in section 2.3, we reported forest floor N mineralization rates relative to total N (rather than total mass), in order to avoid artefact errors resulting from the mixing of mineral soil and organic forest floor. The ratio of mineralizable-to-total N does not inform us on the total amount of N that is available to trees, but serves as an index of soil organic matter quality. Thus, an increase in forest floor N mineralization per total N on scarified plots in Côte-Nord implies that scarification in this climatic region had improved soil nutritional quality, 18 years after treatment. Consistent with this result, there was a 22% decrease in PNUE on scarified plots in Côte-Nord, although this effect was not statistically significant (P = 0.144). Regenerating black spruce forests in cool-humid climates, such as Côte-Nord, are characterized by a dense understory of ericaceous shrubs (Reicis et al. 2020) that reputedly reduce soil N cycling. Ericaceous litter tannin inputs to the forest floor can reduce soil enzyme activities (Joanisse et al. 2007) and the mineralization of organic N (Joanisse et al. 2009). Reicis et al. (2020) reported that canopy gaps allow greater light penetration to the forest floor, which increases the ericaceous shrub cover and ericaceous litter tannin concentrations. From these previous studies, we infer that scarification improves site nutritional quality in Côte-Nord, by accelerating canopy closure thereby reducing light penetration to the forest floor and decreasing the competitive effects of ericaceous shrubs.
Although our study did not focus on the main effects of climatic regions on black spruce physiology, we did observe higher iWUE, PNUE, net rate of photosynthesis (per needle weight) and foliar δ18O in Abitibi than in Côte-Nord. While some of these effects are consistent with expectations (e.g. higher WUE in the warmer-drier climate), others are not (e.g. lower PNUE in the cooler-humid climate). In fact, a simultaneous increase in WUE and PNUE due to drier climate is contrary to the trade-offs predicted by theory (Wright et al. 2003). We deem, however, that it is tenuous to infer mechanisms controlling differences in the physiological status of black spruce trees growing on sites located hundreds of kilometres apart, whereby the contribution of genetic variation may be confounded with that of phenotypic plasticity (Benomar et al. 2016). Furthermore, some of our measured traits, such as δ18O values, are not only influenced by total evapotranspiration, but also by the isotopic composition of soil water, air humidity, precipitation patterns and leaf morphology (Barbour 2007). For these reasons, we believe that it is better to study the physiological responses of black spruce to treatments within given sites, as this is conducive to understanding adaptive mechanisms for future forest management across different regions.
In summary, our study shows that scarification in each climatic region maintains higher relative growth rates of black spruce 18 years after treatment. Scarification increases the total transpiration from trees evenly across the east-to-west moisture gradient in the province of Québec, to which trees adapt by increasing WUE. In the relatively warmer-drier climate, a longer growing season may allow black spruce to benefit from scarification as much as in cooler-humid climate, where scarification is thought to alleviate competition from ericaceous shrubs. On the other hand, scarification appears to improve soil nutritional quality only in the relatively cooler-humid climate.
Acknowledgements
The authors would like to thank D. Guimond and D. Dumais (MFWP) for technical assistance with the LiCor, L. Flanagan (U. of Lethbridge) for his expertise and advice on stable isotopes and F. Rousseau (U. of Sherbrooke) for advice on statistical analyses. We also thank W. Parsons, K. Reicis, T. Audet and S. Cloutier (U. of Sherbrooke) for field and laboratory assistance. The authors thank the technical staff (MFWP) who contributed to establishing and maintaining these experimental sites over time. Financial support was provided by a Natural Sciences and Engineering Research Council of Canada (NSERC) Strategic Project Grant awarded to R.L. Bradley.
References
Barbour M.M. (2007). Stable oxygen isotope composition of plant tissue: a review. Functional Plant Biology 34(2): 83–94. https://doi.org/10.1071/FP06228.
Benomar L., Lamhamedi M.S., Rainville A., Beaulieu J., Bousquet J., Margolis H.A. (2016). Genetic adaptation vs. ecophysiological plasticity of photosynthetic-related traits in young Picea glauca trees along a regional climatic gradient. Frontiers in Plant Science 7: 1–14. https://doi.org/10.3389/fpls.2016.00048.
Blouin J., Berger J.-P. (2002). Guide de reconnaissance des types écologiques de la région écologique 5a – Plaine de l’Abitibi. Ministère des Ressources naturelles du Québec, Forêt Québec, Direction des inventaires forestiers, Division de la classification écologique et productivité des stations. Québec, Québec.
Bradley R.L., Titus B.D., Fyles J.W. (1997). Nitrogen acquisition and competitive ability of Kalmia angustifolia L., paper birch (Betula papyrifera Marsh.) and black spruce (Picea mariana (Mill.) B.S.P.) seedlings grown on different humus forms. Plant and Soil 195: 209–220. https://doi.org/10.1023/A:1004263716346.
Environment Canada (2018). Canadian climate normals 1981–2010. Climate Normals. http://climate.weather.gc.ca. [Cited 20 August 2019].
Farquhar G.D., Lloyd J. (1993). Carbon and oxygen isotope effects in the exchange of carbon dioxide between terrestrial plants and the atmosphere. In: Stable isotopes and plant carbon-water relations. Elsevier. p. 47–70. https://doi.org/10.1016/b978-0-08-091801-3.50011-8.
Farquhar G.D., Barbour M.M., Henry B.K. (1998). Interpretation of oxygen isotope composition of leaf material. In: Griffith H. (ed.). Stable isotopes: integration of biological, ecological and geochemical processes. BIOS Scientific Publishers, Oxford, UK. p. 27–62. https://doi.org/10.1201/9781003076865-3.
Flanagan L.B., Farquhar G.D. (2014). Variation in the carbon and oxygen isotope composition of plant biomass and its relationship to water-use efficiency at the leaf- and ecosystem-scales in a northern Great Plains grassland. Plant, Cell & Environment 37(2): 425–438. https://doi.org/10.1111/pce.12165.
Hamel B., Belanger N., Paré D. (2004). Productivity of black spruce and Jack pine stands in Quebec as related to climate, site biological features and soil properties. Forest Ecology and Management 191(1–3): 239–251. https://doi.org/10.1016/j.foreco.2003.12.004.
Hébert F., Boucher J.F., Walsh D., Tremblay P., Côté D., Lord D. (2014). Black spruce growth and survival in boreal open woodlands 10 years following mechanical site preparation and planting. Forestry 87(2): 277–286. https://doi.org/10.1093/forestry/cpt052.
Hofland-Zijlstra J.D., Berendse F. (2009). The effect of nutrient supply and light intensity on tannins and mycorrhizal colonisation in Dutch heathland ecosystems. Plant Ecology 201(2): 661–675. https://doi.org/10.1007/978-90-481-2798-6_25.
Joanisse G.D., Bradley R.L., Preston C.M., Munson A.D. (2007). Soil enzyme inhibition by condensed litter tannins may drive ecosystem structure and processes: the case of Kalmia angustifolia. New Phytologist 175(3): 535–546. https://doi.org/10.1111/j.1469-8137.2007.02113.x.
Joanisse G.D., Bradley R.L., Preston C.M., Bending G.D. (2009). Sequestration of soil nitrogen as tannin-protein complexes may improve the competitive ability of sheep laurel (Kalmia angustifolia) relative to black spruce (Picea mariana). New Phytologist 181(1): 187–198. https://doi.org/10.1111/j.1469-8137.2008.02622.x.
Johansson M.B. (1994). The influence of soil scarification on the turn-over rate of slash needles and nutrient release. Scandinavian Journal of Forest Research 9(1–4): 170–179. https://doi.org/10.1080/02827589409382828.
Morneau C., Landry Y. (2007). Guide de reconnaissance des types écologiques des régions écologiques 6h – Collines du lac Péribonka et 6i – Hautes collines du réservoir aux Outardes. Québec, Québec. https://mffp.gouv.qc.ca/publications/forets/connaissances/guide-ecologique-6hi.pdf. (Cited 23 March 2019).
Nicault A., Boucher E., Tapsoba D., Arseneault D., Berninger F., Bégin C., DesGranges J., Guiot J., Marion J., Wicha S., Bégin Y. (2015). Spatial analysis of black spruce (Picea mariana (Mill.) B.S.P.) radial growth response to climate in northern Québec – Labrador Peninsula, Canada. Canadian Journal of Forest Research 45(3): 343–352. https://doi.org/10.1139/cjfr-2014-0080.
R Development Core Team (2015). R: a language and environment for statistical computing R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org/.
Rappe George M.O., Hansson L.J., Ring E., Jansson P.E., Gärdenäs A.I. (2017). Nitrogen leaching following clear-cutting and soil scarification at a Scots pine site – a modeling study of a fertilization experiment. Forest Ecology and Management 385: 281–294. https://doi.org/10.1016/j.foreco.2016.11.006.
Reicis K., Bradley R.L., Joanisse G., Houle D., Tremblay S., Barrette M., Wotherspoon A. (2020). Pre-commercial thinning enhances competitive traits of boreal ericaceous shrubs and reduces soil fertility. Forest Ecology and Management 458. https://doi.org/10.1016/j.foreco.2019.117801.
Saucier J.P., Robitaille A., Grondin P. (2009). Cadre bioclimatique du Québec. In: Doucet R., Côté M. (eds.). Manuel de foresterie, 2nd edition. Ordre des ingénieurs forestiers du Québec, Éditions Multimondes, Québec, QC. p. 186–205.
Scheidegger Y., Sauer M., Bahn M., Siegwolf R. (2000). Linking stable oxygen and carbon isotopes with stomatal conductance and photosynthetic capacity: a conceptual model. Oecologia 125: 350–357. https://doi.org/10.1007/s004420000466.
Sikström U., Hjelm K., Hanssen K.H., Saksa T., Wallertz K. (2020). Influence of mechanical site preparation on regeneration success of planted conifers in clearcuts in Fennoscandia – a review. Silva Fennica 54(2) article 10172. https://doi.org/10.14214/sf.10172.
Soil Survey Staff (2006). Keys to soil taxonomy. 10th ed. USDA-Natural Resources Conservation Service, Washington, D.C.
Thiffault N., Jobidon R. (2006). How to shift unproductive Kalmia angustifolia-Rhododendron groenlandicum heath to productive conifer plantation. Canadian Journal of Forest Research 36(10): 2364–2376. https://doi.org/10.1139/x06-090.
Thiffault N., Titus B., Munson A. (2005). Silvicultural options to promote seedling establishment of Kalmia–Vaccinium-dominated sites. Scandinavian Journal of Forest Research 20(2): 110–121. https://doi.org/10.1080/02827580510008356.
Walker X.J., Mack M.C., Johnstone J.F. (2015). Stable carbon isotope analysis reveals widespread drought stress in boreal black spruce forests. Global Change Biology 21(8): 3102–3113. https://doi.org/10.1111/gcb.12893.
Willis J.L., Walters M.B., Gottschalk K.W. (2015). Scarification and gap size have interacting effects on northern temperate seedling establishment. Forest Ecology and Management 347: 237–247. https://doi.org/10.1016/j.foreco.2015.02.026.
Wright I.J., Reich P.B., Westoby M. (2003). Least-cost input mixtures of water and nitrogen for photosynthesis. The American Naturalist 161(1): 98–111. http://bio.mq.edu.au/~iwright/pdfs/wrw03an.pdf. (Cited 4 Oct 2019).
Total of 28 references.
