Kalle Kemppainen  ,
Jari Miina,
Eetu Tarvainen,
Ville Kankaanhuhta,
Juha Laitila,
Heli Peltola,
Markus Strandström,
Kalle Kärhä
,
Jari Miina,
Eetu Tarvainen,
Ville Kankaanhuhta,
Juha Laitila,
Heli Peltola,
Markus Strandström,
Kalle Kärhä
Quality of drill-based site preparation and early performance of Norway spruce and Scots pine seedlings planted in different seasons
Kemppainen K., Miina J., Tarvainen E., Kankaanhuhta V., Laitila J., Peltola H., Strandström M., Kärhä K. (2026). Quality of drill-based site preparation and early performance of Norway spruce and Scots pine seedlings planted in different seasons. Silva Fennica vol. 60 no. 3 article id 26011. https://doi.org/10.14214/sf.26011
Highlights
- A drill-based device for mechanical site preparation significantly reduces soil exposure
- The drill created mostly good (61%) or satisfactory (34%) planting spots for conifer seedlings
- Two years after planting, Norway spruce seedlings performed better than Scots pine seedlings.
Abstract
Mechanical site preparation (MSP) is essential for successful forest regeneration. However, excessively exposed soil may harm the environment and increase the total costs of seedling stand management. A drill-based MSP device (drill), designed to reduce soil exposure, was recently tested in Finland. This study investigated the quality of drill-based MSP and the early performance of Norway spruce (Picea abies (L.) Karst.) and Scots pine (Pinus sylvestris L.) seedlings planted in different seasons on five mineral soil stands. The findings showed that the mean area of the drilling spots (0.18 m2) was about 20% of that reported for mounding. The drill mostly resulted in good (61%) or satisfactory (34%) planting spot quality (e.g., good spots were covered with mineral soil). Ground and soil obstacles, such as roots, stones, stumps, and logging residues, negatively affected planting spot quality. About 13% of the planting spots did not have a pure mineral soil cover, which is required to mitigate pine weevil (Hylobius abietis L.) damage. Most seedlings were still alive one year after planting, but the proportion of healthy seedlings was 71% for Norway spruce and 48% for Scots pine two years after planting. Overall, drill-based MSP may be most feasible for drier sites with less competitive ground vegetation. However, the productivity and costs, the performance of the planted seedlings, and other potential benefits of drill-based MSP should be further tested under varying operating conditions and compared with those of conventional MSP methods.
Keywords
forest regeneration;
seedling survival;
manual planting;
mechanical soil preparation
-
Kemppainen,
School of Forest Sciences, University of Eastern Finland, Yliopistokatu 7, FI-80100 Joensuu, Finland; Natural Resources Institute Finland, Yliopistokatu 6, FI-80100 Joensuu, Finland
 https://orcid.org/0009-0000-6184-8812
E-mail
kalle.kemppainen@uef.fi
https://orcid.org/0009-0000-6184-8812
E-mail
kalle.kemppainen@uef.fi
-
Miina,
Natural Resources Institute Finland, Yliopistokatu 6, FI-80100 Joensuu, Finland
https://orcid.org/0000-0002-8639-4383
E-mail
jari.miina@luke.fi
- Tarvainen, School of Forest Sciences, University of Eastern Finland, Yliopistokatu 7, FI-80100 Joensuu, Finland E-mail eetu.tarvainen@harvestia.fi
-
Kankaanhuhta,
Natural Resources Institute Finland, Yliopistokatu 6, FI-80100 Joensuu, Finland
https://orcid.org/0000-0001-5785-5972
E-mail
ville.kankaanhuhta@luke.fi
-
Laitila,
Natural Resources Institute Finland, Yliopistokatu 6, FI-80100 Joensuu, Finland
https://orcid.org/0000-0003-4431-3319
E-mail
juha.laitila@luke.fi
-
Peltola,
Natural Resources Institute Finland, Yliopistokatu 6, FI-80100 Joensuu, Finland
https://orcid.org/0000-0003-1384-9153
E-mail
heli.peltola@uef.fi
-
Strandström,
Metsäteho Oy, Vernissakatu 1, FI-01300 Vantaa, Finland
https://orcid.org/0009-0004-0868-3042
E-mail
markus.strandstrom@metsateho.fi
-
Kärhä,
School of Forest Sciences, University of Eastern Finland, Yliopistokatu 7, FI-80100 Joensuu, Finland
https://orcid.org/0000-0002-8455-2974
E-mail
kalle.karha@uef.fi
Received 6 February 2026 Accepted 23 June 2026 Published 1 July 2026
Views 4206
Available at https://doi.org/10.14214/sf.26011 | Download PDF

Supplementary Files
1 Introduction
Manual planting of conifer seedlings following mechanical site preparation (MSP) is among the most common forest regeneration methods in Nordic boreal forests (Ramantswana et al. 2020; Sikström et al. 2020; Natural Resources Institute Finland 2025; Skogsstyrelsen 2025). MSP is an essential part of forest regeneration because it ensures favorable soil temperature, moisture, and nutrient conditions for planted seedlings (Örlander et al. 1990; Sutton 1993; Heiskanen et al. 2019), thereby improving their survival and growth (Hjelm et al. 2019; Sikström et al. 2020; Uotila et al. 2022). An appropriate MSP method should be selected according to the characteristics of each site, including soil properties and site fertility (Heiskanen et al. 2019; Sikström et al. 2020). The most suitable MSP practices also depend on the intended forest regeneration method and tree species (Luoranen et al. 2007).
MSP methods are commonly grouped into mound-forming techniques (mounding) and those that only remove organic layers, usually a mor humus layer (humus layer) in boreal forests, to expose mineral soil (patch scarification and disc trenching) (Örlander et al. 1990; Sutton 1993; Luoranen et al. 2007). In the context of planting, however, all MSP methods aim to prepare a pure mineral soil cover (at least 15–20 cm) around planted seedlings, which efficiently prevents pine weevil (Hylobius abietis L.) damage (Örlander et al. 1990; Björklund et al. 2003; Nordlander et al. 2005).
Mounding inverts the humus layer so that the bare mineral soil is placed on top of the mound, whereas the lower part of the mound consists of a single (inverting) or double (ditch and spot mounding) humus layer (Sikström et al. 2020). Mounding is typically used for planting Norway spruce (Picea abies (L.) Karst.) on mesic and herb-rich heath sites with fine- or medium-coarse-textured mineral soils, and can also be used for planting Scots pine (Pinus sylvestris L.) on mesic sites with coarse-textured mineral soils and sub-xeric sites with fine-textured mineral soils (Luoranen et al. 2007; Äijälä et al. 2019). Mounds enhance seedling growth by increasing soil temperature and improving nutrient availability (Örlander et al. 1998; Heiskanen and Rikala 2006; Smolander and Heiskanen 2007) and by reducing competition from ground vegetation (Örlander et al. 1996; Nilsson and Örlander 1999; Luoranen et al. 2007).
Scarification techniques, such as patch scarification and disc trenching, are most suitable for sub-xeric or xeric sites with medium- or coarse-textured mineral soils, where natural regeneration (with seed trees) or seeding of Scots pine is more common than planting (Luoranen et al. 2007; Äijälä et al. 2019). Disc trenching differs from mounding and patch scarification by creating continuous furrows rather than directed planting spots (Sikström et al. 2020).
In addition to a suitable MSP method, high-quality planting spots and properly executed planting are vital for successful forest regeneration (Dumins and Lazdina 2018; Pikkarainen et al. 2020; Luoranen et al. 2021). The use of continuously advancing mounders may be limited, as stony soil and fresh logging residues decrease planting spot quality (Saksa et al. 2018). Moreover, planting season and weather conditions may significantly affect seedling performance for different tree species. For example, late summer plantings of Scots pine seedlings have failed more often than early-summer plantings (Pikkarainen et al. 2020). Furthermore, warm and dry summers can particularly increase the mortality of planted Norway spruce seedlings (Nordin et al. 2022; Luoranen et al. 2023).
Although MSP clearly improves forest regeneration, it may also have adverse effects. In some cases, widely used MSP methods can excessively disturb the soil surface, with environmental, social, and economic implications. The average proportion of disturbed soil can account for 52% and 37% of the forest regeneration area in disc trenching and mounding, respectively (Sikström et al. 2020). Consequently, clear cutting combined with intensive MSP may lead to erosion and nutrient leaching (Alcázar et al. 2002; Piirainen et al. 2009; Palviainen et al. 2014) or decrease landscape values (Karjalainen 2006). Moreover, conflicts among different stakeholders are especially common in regions where reindeer husbandry is practiced alongside forestry (Ersson et al. 2023) since MSP reduces ground vegetation, including important reindeer forage lichens (Ring and Sikström 2024). Nevertheless, prepared soil is a favorable substrate for pioneer species, such as naturally regenerated broadleaf trees (Laine et al. 2020; Lidman et al. 2024). Therefore, excessive soil exposure may reduce seedling growth and increase the costs of seedling stand management (Uotila et al. 2010).
Overall, reducing adverse effects without compromising the success of forest regeneration requires novel MSP technologies and practices (Ring and Sikström 2024; Thiffault et al. 2025). A good premise for such innovations is the preference for intermittent and directed MSP methods (Löf et al. 2012). Among the first in the Nordic countries, Hansson et al. (2024) tested the concept of an autonomous mechanized tree seedling planting machine in Sweden. Aiming to develop a machine that reduces soil disturbance yet produces a suitable planting spot, they found a drill-based MSP device (drill) to be the most promising of three diverse concepts. However, different manual, motor-manual, and mechanized drills are already used for pitting in plantation forestry (McEwan and Steenkamp 2014; Hechter et al. 2020; Zulu et al. 2024). In Finland, a novel drill that creates circular planting spots was also recently developed by the forest machine manufacturer Ponsse Plc. Although the drill is expected to reduce soil exposure compared with conventional MSP methods, its performance in practical forestry has not yet been thoroughly investigated.
This study investigated the quality of drill-based MSP and the early performance of Norway spruce and Scots pine seedlings planted in different seasons on five mineral soil stands. Planting spots were classified as poor, satisfactory, or good based on their dimensions and mineral soil cover. The main factors reducing planting spot quality, such as stones and humus layer residues within the planting spots, were also recorded. To determine the effect of season on planting spot quality and early seedling performance, drill-based MSP and planting were conducted in early, mid, and late summer for Norway spruce and in early and late summer for Scots pine. Early performance was evaluated based on the number and height of healthy, weakened, and dead seedlings during two growing seasons after planting.
2 Materials and methods
2.1 Study sites, machinery, and manual planting
Five experimental stands were established in state-owned forest regeneration areas in eastern Finland (Fig. 1), where logging residues and stumps were left on sites after clear cutting. All sites underwent MSP using a drill with a 40-cm diameter (Fig. 1) mounted on a Volvo EC 219 CL crawler excavator (operating weight: 20 t; engine power: 123 kW). The drill was developed and built by Ponsse Plc (Vieremä, Finland). The drilling method is most comparable to patch scarification, as both create planting spots slightly below the ground surface (5–10 cm) by removing the humus layer above the mineral soil (Luoranen et al. 2007). The drill occasionally failed to move the entire humus layer outside the planting spot. Consequently, humus layer residues were in some cases partially mixed with mineral soil or left on top of the planting spots. The drill’s rotational speed was relatively slow (around 70–90 r min−1), and it did not require much pressure from the boom to prepare the soil. The drill needed to rotate several times for each planting spot, so the estimated time per spot was approximately 4–8 s. However, this study did not examine productivity in greater detail.
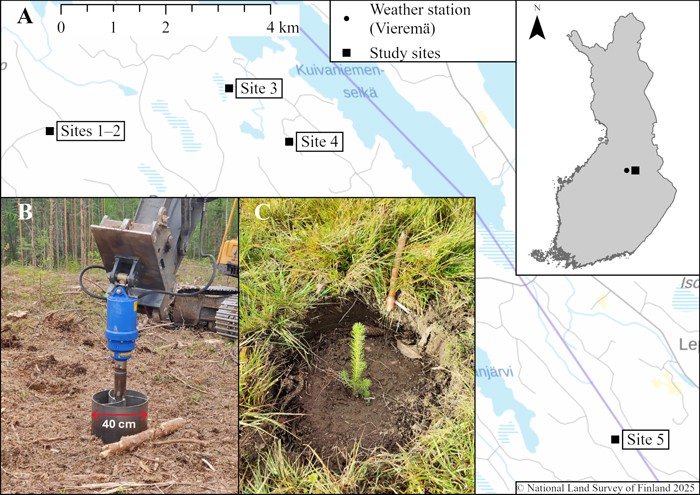
Fig. 1. In 2022, study sites (A) were established in eastern Finland (27°53′58″–28°06′58″ E, 63°44′25″–63°48′06″ N). The drill diameter was 40 cm (B). Sites 1 and 2 were planted with Norway spruce seedlings, and sites 3–5 were planted with Scots pine seedlings. Panel (C) shows a Norway spruce seedling planted in late summer. The map was created using ArcGIS Pro (version 3.1.0). Photos (B and C): Eetu Tarvainen.
To examine the effect of drilling and planting season on planting spot quality and early performance of seedlings, a machine operator with 20 years of experience prepared a total of 4120 planting spots in three separate periods in 2022: early (week 22, n = 1642), mid (week 29, n = 797), and late (weeks 35 and 36, n = 1681) summer. Norway spruce seedlings were planted throughout the growing season (Table 1). In contrast, Scots pine seedlings were planted only in early and late summer, as midsummer planting is not recommended for Scots pine (Äijälä et al. 2019) and suitable seedlings are not usually available at that time. Seedlings were generally planted immediately after MSP to ensure the planting spots were fresh. However, the delay between MSP and planting extended to four weeks for the late summer Norway spruce plantings at site 2. Following Finnish forest management recommendations, the target planting density was 1800 seedlings ha−1 for Norway spruce and 2000–2200 seedlings ha−1 for Scots pine (Äijälä et al. 2019). In practice, one seedling was planted at each spot, so the actual planting density varied according to how many planting spots were prepared.
Norway spruce and Scots pine seedlings were delivered from different nurseries to the study sites in open plastic trays. The seedlings planted in late summer came from different growing batches than those planted in early or midsummer, which caused the difference in initial seedling heights. Specifically, 1-year-old container seedlings (all Scots pine and late summer Norway spruce seedlings) and 1.5-year-old container seedlings (early and midsummer Norway spruce seedlings) were planted by professional forest workers. All planted seedlings were treated in the nursery with a pesticide against pine weevils (Karate Zeon or Hallmark Zeon).
| Table 1. Site details for Norway spruce and Scots pine seedlings planted in different seasons during the summer of 2022. In the first measurement (2022), all planting spots were measured (n = 4120). The follow-up measurements in 2023 and 2024 were based on 60 permanent circular sample plots (r = 5.64 m). Planting season: early (ES), mid (MS), and late (LS) summer. Growing season: the number of growing seasons at the end of the study period (2024). Site type: herb-rich heath (Oxalis-Myrtillus type [OMT]), mesic heath (Vaccinium myrtillus type [MT]), and sub-xeric heath (Vaccinium vitis-idaea type [VT]) forests. | ||||||||
| Area (ha) | Spots and seedlings | Sample plots | Growing seasons | Site type | ||||
| Site | Tree species/Season | 2022 | 2023 | 2024 | ||||
| 1 | Norway spruce | 0.6 | 1251 | 376 | 337 | 18 | ||
| ES | 0.2 | 414 | 102 | 82 | 6 | 2.0 | MT | |
| MS | 0.2 | 393 | 125 | 114 | 6 | 1.5 | MT | |
| LS | 0.2 | 444 | 149 | 141 | 6 | 1.0 | MT | |
| 2 | Norway spruce | 0.6 | 1250 | 374 | 279 | 18 | ||
| ES | 0.2 | 406 | 125 | 92 | 6 | 2.0 | OMT | |
| MS | 0.2 | 404 | 118 | 98 | 6 | 1.5 | OMT | |
| LS | 0.2 | 440 | 131 | 89 | 6 | 1.0 | OMT | |
| 3 | Scots pine | 0.2 | 400 | 111 | 78 | 6 | ||
| ES | 0.2 | 400 | 111 | 78 | 6 | 2.0 | VT | |
| 4 | Scots pine | 0.2 | 422 | 124 | 74 | 6 | ||
| ES | 0.2 | 422 | 124 | 74 | 6 | 2.0 | VT | |
| 5 | Scots pine | 0.4 | 797 | 289 | 221 | 12 | ||
| LS | 0.4 | 797 | 289 | 221 | 12 | 1.0 | VT | |
| Total | 2.0 | 4120 | 1274 | 989 | 60 | |||
Each study site was defined using Cajander’s (1926) site classification (site type), which is based on the composition of ground vegetation species (Table 1). Scots pine seedlings were planted in sub-xeric heath forest (Vaccinium vitis-idaea type [VT]). In contrast, Norway spruce seedlings were planted in mesic heath forest (Vaccinium myrtillus type [MT]) and herb-rich heath forest (Oxalis-Myrtillus type [OMT]). All study sites had medium-textured mineral soil with a grain size of 0.06–0.6 mm (Luoranen et al. 2007). Although Scots pine planting sites were slightly rockier than Norway spruce sites, some Scots pine planting areas at site 5 were paludified, leading to approximately 3% of the planting spots being covered with peat rather than mineral soil.
2.2 Field measurements
The initial measurement in 2022 focused on planting spot quality, which was visually classified as good, satisfactory, or poor for all created planting spots (n = 4120, Supplementary file S1). Quality was considered good when the mineral soil cover around the planted seedling was approximately 20 cm, and the spot was otherwise suitable for planting (e.g., free of stones); satisfactory when factors reducing quality, such as stones and roots within the planting spot, were present and/or the mineral soil cover was markedly less than 20 cm; and poor when several factors reducing quality and hindering planting, including the residues of humus layer, stones, roots, and/or logging residues, were present. In addition, the diameter and depth (cm) of each planting spot were measured from the top of the humus layer.
Follow-up measurements were conducted in late spring 2023 and 2024 before seedlings started to grow. These inventories focused on measuring the height and evaluating the vitality of planted seedlings using 60 circular sample plots (r = 5.64 m) that were systematically placed at the study sites (Table 1, Suppl. file S2). In both follow-up measurements, the same permanent sample plots and variables were recorded. Seedlings and planting spots were not tracked individually during these measurements.
In the follow-up measurements, seedlings were classified as healthy, weakened, or dead (Suppl. file S1) and as planted in the middle, side, or outside the planting spot (seedling position). The planting depth of each seedling was classified as proper (2–3 cm), shallow (root plug shallow and/or exposed), or deep (shoot clearly less visible than usual). Seedling stem angles were classified as upright (<20°), askew (20°–40°), or prone (>40°). Finally, soil compaction around the planted seedlings was assessed by gently pulling them; whenever they seemed easily detachable from the soil, they were classified as uncompacted. In addition to the seedling measurements, spot diameters were remeasured, and the spot bottoms below the ground surface were classified as hole, flat, or elevated (spot type). The last spot quality classification occurred in 2023, as ground vegetation hindered evaluation accuracy in 2024.
The additional spot- and seedling-specific notes documented for each measurement are presented in Table 2, including observations of potential negative factors affecting the quality of planting spots, such as roots, stumps, stones, and logging residues. In addition, planting spots with no bare mineral soil cover were marked to contain humus layer residues. While remarks on peat denoted planting spots in paludified areas, any presence of water within the planting spots was also recorded separately. Brushwood was defined as naturally regenerated broadleaf trees that may compete with planted seedlings. Notes on logging trails referred to machine tracks formed during the harvesting. In addition, tree-top browsing and breakages, and seedlings that had come out of the soil (loose), were recorded.
In 2022, notes on humus layer residues (13%), stones (9%), and roots (5%) were common (Table 2). The number of brushwood notes increased between 2022 and 2024, indicating that naturally regenerated broadleaf trees grew during the study period. Seedling damage was relatively rare in 2023 and 2024 (Table 2).
| Table 2. Spot- and seedling-specific notes that were documented in 2022, 2023, and 2024. Notes that were not recorded in 2022 are marked with “-”. In the first measurement, all planting spots were measured and evaluated (n = 4120). The follow-up measurements in 2023 (n = 1274) and 2024 (n = 989) were based on 60 permanent circular sample plots (r = 5.64 m) systematically placed at the study sites. | ||||||
| Note | Measurement year | |||||
| 2022 | 2023 | 2024 | ||||
| n | % | n | % | n | % | |
| No notes | 2864 | 70 | 408 | 32 | 351 | 35 |
| Stumpa | 24 | 1 | 12 | 1 | 11 | 1 |
| Roota | 203 | 5 | 221 | 17 | 83 | 8 |
| Stonea | 351 | 9 | 158 | 12 | 183 | 19 |
| Logging residuesa | 95 | 2 | 59 | 5 | 57 | 6 |
| Humus layer residuesb | 518 | 13 | 267 | 21 | 1 | 0 |
| Waterc | 53 | 1 | 46 | 4 | 22 | 2 |
| Logging traild | 12 | 0 | 1 | 0 | 2 | 0 |
| Peate | - | - | 40 | 3 | 0 | 0 |
| Brushwoodf | - | - | 1 | 0 | 273 | 28 |
| Tree-top browsingg | - | - | 52 | 4 | 6 | 1 |
| Tree-top breakageg | - | - | 2 | 0 | 0 | 0 |
| Loose seedlingg | - | - | 7 | 1 | 0 | 0 |
| Total | 4120 | 100 | 1274 | 100 | 989 | 100 |
| a Ground and soil obstacles reducing planting spot quality b Humus layer residues within the planting spot c The presence of water within the planting spot d Machine tracks formed during the harvesting e Paludified areas where planting spots were covered with peat f Naturally regenerated broadleaf trees g Seedling damage affecting vitality | ||||||
The same forest worker conducted the field inventories in 2022 and 2023, while two different forest workers conducted the final follow-up measurements in 2024. After the measurements, the data were checked, and any errors were corrected. Only eight observations with partially missing data needed to be excluded from the final dataset (Table 1), which consisted of 4120 planting spots for 2022 (all prepared spots), 1274 planting spots and seedlings for 2023 (based on 60 permanent sample plots), and 989 planting spots and seedlings for 2024 (based on 60 permanent sample plots).
2.3 Weather conditions
The weather data were obtained from the nearest meteorological station in Vieremä (63.8° N, 27.2° E) (Fig. 1) (Finnish Meteorological Institute 2024). Information about the monthly minimum, maximum, and mean temperatures (°C) and the effective temperature sum, along with precipitation (mm), during the growing seasons in 2022 and 2023 is provided in Suppl. file S3. The effective temperature sum was calculated with a threshold of +5 °C. The long-term weather conditions from 1990 to 2021 were also assessed to determine whether weather conditions during the study period were normal or unusual. The weather data for 2024 were excluded because the sites had already been measured at the beginning of June.
When forest regeneration operations were conducted in 2022, the weather was warmer and drier than average. The summer of 2022 was particularly hot, but in September, the daily temperatures were lower than typical. The weather conditions in 2023 were more variable than normal: the growing season was notably wetter but also warmer; the effective temperature sum was 17% higher than the long-term average from 1990 to 2021; and the minimum observed air temperature was −2 °C in June, much lower than the 4 °C in 2022 or the average of 3 °C from 1990 to 2021.
2.4 Statistical analysis
The ordinal response variables (planting spot quality and seedling vitality) were analyzed using cumulative link models (CLMs). The probabilities of planting spots having poor (j = 1), satisfactory (j = 2), or good (j = 3) quality were estimated using a CLM, which evaluated the cumulative probabilities of an ordinal response belonging to a class j or lower on a logit scale (Agresti 2010). Under the proportional odds assumption, the regression parameters or thresholds of CLMs cannot vary across response categories or depend on regression parameters (Christensen 2026a). The CLM also included variables used to classify planting spot quality as predictors to control for their effects on planting spot quality and to determine the effects of site type, season, and spot dimensions. Consideration of random effects was unnecessary in the spot quality analysis, as all planting spots (n = 4120) across five study sites were evaluated, yielding a limited number of groups (n = 5) and a simple two-level hierarchical structure. Moreover, the CLM with site-level random effects did not converge and showed worse model fit than the simpler CLM without random effects.
The data on seedling vitality, collected in 2023 and 2024, consisted of three nested observation levels (Table 1): planted seedlings (n = 2263), sample plots (n = 60), and study sites (n = 5). Therefore, seedling vitality (1 = dead, 2 = weakened, or 3 = healthy) was modeled using a cumulative link mixed model (CLMM) with sample plot as a random effect, as the same sample plots were remeasured in 2023 and 2024. The explanatory variables included interaction terms among site types, planting seasons, and tree species, as well as notes on factors that may influence vitality (e.g., seedling damage). The following CLMM with the proportional odds assumption was used (Christensen 2026a, 2026b):
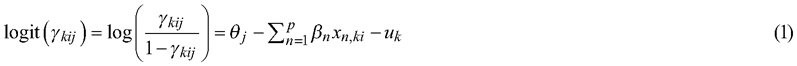
where γkij is the cumulative probability that the ith observation of the kth sample plot falls in the jth category or below (j = 1, 2, 3), θj is the threshold (i.e., cutoff point) for the jth category, βn is the fixed coefficient of explanatory variable xn (n = 1, 2, …, p), and uk is a random effect for the kth sample plot. Note that the CLM fitted for planting spot quality only differed from Eq. 1 in that no sample plots were measured, and thus random effects were excluded from the linear predictor.
Only the height development of Norway spruce seedlings was statistically analyzed, as the Scots pine seedlings planted in early and late summer were from different growing batches, resulting in differences in their initial heights. In addition, differences between site types could not be evaluated for Scots pine, as all seedlings were planted on sub-xeric heath forest (VT) sites (Table 1).
The following linear mixed-effect model (LMEM) was fitted for Norway spruce seedling height (Mehtätalo and Lappi 2020):
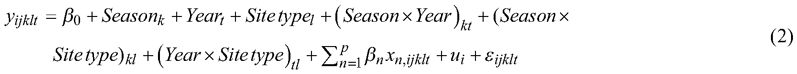
where the response variable y is the height (cm) of seedling j on sample plot i, Season is the planting season (k = early, mid, late), Year is the measurement year (t = 2023, 2024), Site type is the site type (l = OMT, MT), β0 is the fixed intercept, βn (n = 1, 2, 3, …, p) are fixed coefficients of explanatory variables xn (seedling vitality, position, planting depth, stem angle, surrounding soil compaction, and notes) representing the seedling effect, ui is a random effect for the ith sample plot, and εijklt is the random error. The three-way interaction among season, year, and site type was not included, simplifying Eq. 2. Note that the seedlings in the sample plots were not tracked individually during the 2023 and 2024 measurements. This analysis excluded seedlings that were dead, had a broken or eaten top, or were loose from the soil (n = 1343).
All statistical analyses were performed using R (version 4.4.3) (R Core Team 2025), and p-values < 0.05 were considered statistically significant. CLMs and CLMMs were fitted using the clm and clmm2 functions from the ordinal package, respectively (Christensen 2025). LMEMs were fitted using the lme function from the nlme package with a power variance function to account for the heteroscedasticity of the residuals (i.e., allowing the residual variance to be proportional to a power of the fitted values) (Mehtätalo and Lappi 2020; Pinheiro and Bates 2026). CLM parameters were estimated using the maximum likelihood method, and LMEM parameters were estimated using the restricted maximum likelihood method. Model superiority was assessed using the Akaike information criterion and by comparing nested models using the likelihood ratio test.
3 Results
3.1 Planting spot dimensions and quality
The mean depth across the planting spots was 10.4 ± 4.5 cm. Planting spots were slightly deeper on the sites planted with Scots pine seedlings (Table 3). The mean diameter across planting spots was 47 ± 6.8 cm. The mean area was greater among planting spots prepared in late summer than among those prepared in early summer (Table 3). Based on the mean planting spot areas, the proportion of exposed soil from the total MSP area ranged from 2.8% to 4.8%, with a target density of 2000 planting spots ha−1.
| Table 3. Details of planting spots (depth, diameter, and area) by season (early [ES], mid [MS], and late [LS] summer) for Norway spruce and Scots pine planting sites (n = 4120). Data are presented as the mean ± standard deviation. | ||||
| Season/ | Planting spot | |||
| Tree species | Depth (cm) | Diameter (cm) | Area (cm2) | Number |
| ES | 10.1 ± 4.4 | 41.9 ± 3.8 | 1388 ± 255 | 1642 |
| Norway spruce | 8.9 ± 3.8 | 42.0 ± 3.4 | 1393 ± 210 | 820 |
| Scots pine | 11.3 ± 4.5 | 41.8 ± 4.2 | 1384 ± 293 | 822 |
| MS | 9.8 ± 4.6 | 46.8 ± 4.8 | 1735 ± 359 | 797 |
| Norway spruce | 9.8 ± 4.6 | 46.8 ± 4.8 | 1735 ± 359 | 797 |
| Scots pine | - | - | - | - |
| LS | 10.9 ± 4.6 | 52.0 ± 6.2 | 2156 ± 539 | 1681 |
| Norway spruce | 8.9 ± 3.4 | 49.8 ± 4.2 | 1960 ± 321 | 884 |
| Scots pine | 13.2 ± 4.6 | 54.5 ± 7.0 | 2373 ± 640 | 797 |
| All | 10.4 ± 4.5 | 47.0 ± 6.8 | 1768 ± 537 | 4120 |
| Norway spruce | 9.2 ± 4.0 | 46.3 ± 5.3 | 1702 ± 384 | 2501 |
| Scots pine | 12.2 ± 4.7 | 48.0 ± 8.6 | 1871 ± 700 | 1619 |
Among the planting spots examined, 61% were classified as good overall, varying from 55% to 68% across sites and seasons (Fig. 2), while 34% were classified as satisfactory and 5% as poor. The type of planting spot was mainly recorded as either flat (49%) or hole (47%).
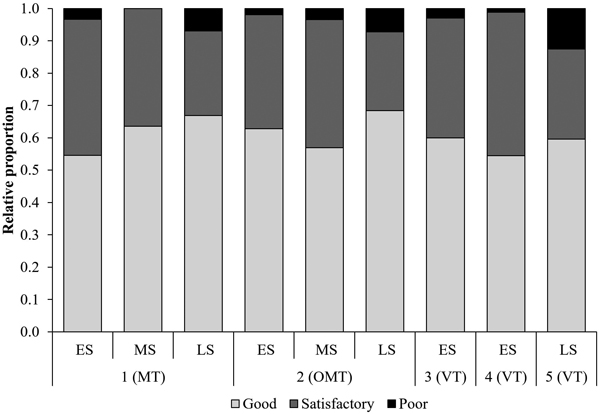
Fig. 2. Proportions of planting spots classified as good, satisfactory, and poor by study site and season (n = 4120). Season: early (ES), mid (MS), and late (LS) summer. Site type: herb-rich heath (Oxalis-Myrtillus type [OMT]), mesic heath (Vaccinium myrtillus type [MT]), and sub-xeric heath (Vaccinium vitis-idaea type [VT]) forests.
The CLM for planting spot quality included a site type × season interaction term, continuous variables (spot diameter and depth), and notes describing factors decreasing quality (e.g., stones, roots, and humus layer residues). These variables negatively or positively influenced the probability that planting spots belong to each quality class (poor, satisfactory, or good); a negative coefficient indicates a decreased probability of belonging to a higher quality class (Table 4).
According to the CLM, the odds of poor planting spot quality were highest for drill-based MSP in early summer (Table 4), and the odds of good planting spot quality were highest for drill-based MSP in midsummer and late summer at MT and OMT sites. Therefore, planting spots for Norway spruce (MT and OMT) had greater odds of belonging to a higher quality class than those for Scots pine (VT). The CLM also indicated that increasing spot depth or diameter was associated with lower odds of belonging to a lower quality class. However, as spot depth increased by 10–20 cm from ground level, the positive effect began to diminish, as the effect of spot depth followed a second-order polynomial, which, in turn, reduced the odds of belonging to a higher quality class. The odds of belonging to the good-quality class were reduced in the following order by the dummy variables (notes): water, logging residues, organic matter, stones, and stumps or roots (Table 4). In the 2022 data, the quality of 26% of the planting spots decreased due to these factors. Nevertheless, 13% of the planting spots were classified as satisfactory or poor with no recorded notes. The CLM accurately predicted the good-quality class (95%), but it predicted the lower quality classes less accurately (Suppl. file S4). The overall accuracy was 79%.
| Table 4. Cumulative link model (CLM) for the quality classification of the planting spots based on data from 2022. The CLM predicts the probability of belonging to the quality classes poor, satisfactory, or good. The threshold coefficients determine the cutoffs between quality classes (poor-to-satisfactory and satisfactory-to-good). The cumulative odds ratios (CORs) are shown in the last column. Site type × Season: interaction coefficients between site types and planting seasons. Site type: herb-rich heath (Oxalis-Myrtillus type [OMT]), mesic heath (Vaccinium myrtillus type [MT]), and sub-xeric heath (Vaccinium vitis-idaea type [VT]) forests. Season: early (ES), mid (MS), and late (LS) summer. | ||||
| Parameter | Estimate | Std. error | p-value | COR |
| Spot diameter | 0.023 | 0.008 | 0.007 | 1.023 |
| Spot depth | 0.134 | 0.021 | <0.001 | 1.144 |
| Spot depth2/100 | −0.333 | 0.076 | <0.001 | 0.716 |
| Notes (ref. No notes) | ||||
| Stump or roota | −3.168 | 0.166 | <0.001 | 0.042 |
| Stonea | −3.293 | 0.156 | <0.001 | 0.037 |
| Logging residuesa | −4.959 | 0.261 | <0.001 | 0.007 |
| Waterb | −7.134 | 0.350 | <0.001 | 0.001 |
| Humus layer residuesc | −4.609 | 0.150 | <0.001 | 0.010 |
| Site type × Season (ref. MT × ES) | ||||
| MT × MS | 1.974 | 0.197 | <0.001 | 7.199 |
| MT × LS | 1.451 | 0.195 | <0.001 | 4.266 |
| OMT × ES | 0.433 | 0.161 | 0.007 | 1.541 |
| OMT × MS | 1.402 | 0.192 | <0.001 | 4.064 |
| OMT × LS | 1.231 | 0.193 | <0.001 | 3.425 |
| VT × ES | −0.257 | 0.135 | 0.057 | 0.773 |
| VT × LS | 0.834 | 0.190 | <0.001 | 2.303 |
| Threshold coefficients | ||||
| Poor | Satisfactory | −3.065 | 0.398 | ||
| Satisfactory | Good | 1.073 | 0.390 | ||
| a Ground and soil obstacles reducing planting spot quality b The presence of water within the planting spot c Humus layer residues within the planting spot | ||||
3.2 Quality of manual planting
The soil around the planted seedlings was typically compacted, and seedlings were mainly planted at a proper depth (Fig. 3). The stem angle of the planted seedlings was usually upright, even though around 30% of the seedlings were slightly askew. Although almost all seedlings were planted at the prepared planting spots, their positions within them varied (Fig. 3).
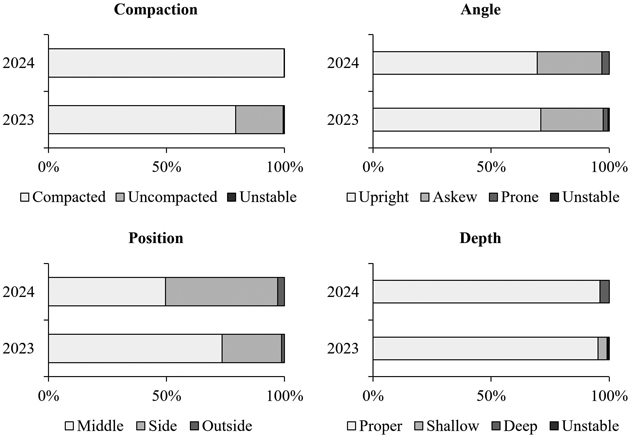
Fig. 3. Soil compaction around the planted seedlings (compacted or uncompacted), seedling stem angle (upright [<20°], askew [20°–40°], or prone [>40°]), seedling position (planted in the middle, side, or outside of the planting spot), and planting depth (proper [2–3 cm], shallow [root plug shallow and/or exposed], or deep [shoot clearly less visible than usual]) based on follow-up measurements in 2023 (n = 1274) and 2024 (n = 989). Unstable refers to seedlings that were loose from the soil.
3.3 Proportion and number of healthy seedlings
The proportion and number of healthy seedlings varied considerably among tree species, site types, planting seasons, and sites (Figs. 4 and 5). In 2023, 86% of Norway spruce seedlings and 60% of Scots pine seedlings were classified as healthy. However, the number of measured seedlings decreased by 2024, particularly at Scots pine planting sites (Fig. 4). As missing seedlings were considered in the 2024 calculation, 71% of Norway spruce seedlings and 48% of Scots pine seedlings were classified as healthy that year. The mean number of healthy seedlings was 1794 and 1478 seedlings ha−1 for Norway spruce and 1313 and 1054 seedlings ha−1 for Scots pine in 2023 and 2024, respectively.
Although the proportion of healthy Norway spruce seedlings was lowest among those planted in late summer at the OMT site, Norway spruce and Scots pine seedlings planted in late summer generally performed better than those planted in early summer. In addition, Norway spruce seedlings planted in midsummer were more successful than those planted in early summer in 2024. Scots pine seedlings planted in early summer had the lowest proportion and number of healthy seedlings in the entire dataset (Figs. 4 and 5).
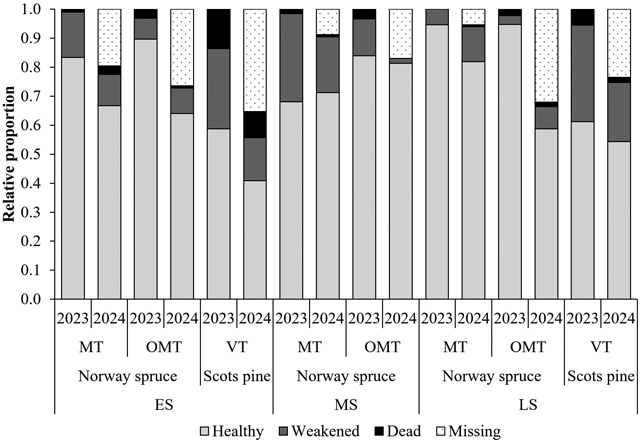
Fig. 4. Observed seedling vitality (healthy, weakened, or dead) by planting season, tree species, site type, and measurement year (2023: n = 1274 and 2024: n = 989). The number of missing seedlings (n = 285) was calculated by subtracting the measured number of seedlings in 2024 from the measured number of seedlings in 2023. Planting season: early (ES), mid (MS), and late (LS) summer. Site type: herb-rich heath (Oxalis-Myrtillus type [OMT]), mesic heath (Vaccinium myrtillus type [MT]), and sub-xeric heath (Vaccinium vitis-idaea type [VT]) forests.
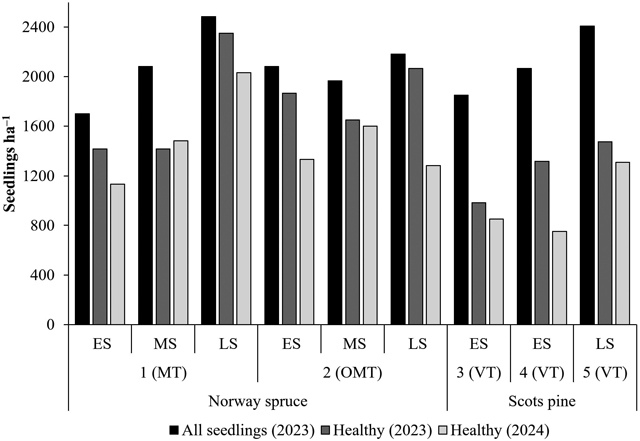
Fig. 5. Seedling number (all seedlings) and number of healthy seedlings per hectare based on the measured permanent sample plots in 2023 and 2024. Planting season: early (ES), mid (MS), and late (LS) summer. Site type: herb-rich heath (Oxalis-Myrtillus type [OMT]), mesic heath (Vaccinium myrtillus type [MT]), and sub-xeric heath (Vaccinium vitis-idaea type [VT]) forests.
According to the CLMM, Norway spruce seedlings were more likely to be healthy than Scots pine seedlings (Table 5). Nevertheless, over half of the individual interaction coefficients among tree species, site types, and measurement years were not significant, although the interaction term was significant overall (p < 0.05). The CLMM predicted that increasing stem skewness was associated with greater odds of belonging to a lower vitality class. In addition, poor soil compaction around the planted seedlings, notes on seedlings with eaten tops, and the presence of water in planting spots decreased the odds of belonging to the healthy vitality class. The variance of the random effect (sample plot) was relatively small (Table 5). The CLMM with fixed variables demonstrated excellent accuracy in predicting the healthy vitality class, although it mostly failed to predict weakened and dead vitality classes (Suppl. file S4). The influence of planting spot quality on seedling vitality was also examined by fitting a CLMM to the follow-up data from 2023, but it was not a significant predictor.
| Table 5. Cumulative link mixed model (CLMM) for the vitality classification of the planted seedlings based on data collected in 2023 and 2024. The CLMM predicts the probability of belonging to the seedling vitality class dead, weakened, or healthy with the sample plot-level random effect (variance). The threshold coefficients determine the cutoffs between vitality classes (dead-to-weakened and weakened-to-healthy). The cumulative odds ratios (CORs) are shown in the last column. Tree species × Site type × Season × Year: interaction coefficients among tree species, site types, planting seasons, and measurement years. Site type: herb-rich heath (Oxalis-Myrtillus type [OMT]), mesic heath (Vaccinium myrtillus type [MT]), and sub-xeric heath (Vaccinium vitis-idaea type [VT]) forests. Planting season: early (ES), mid (MS), and late (LS) summer. | ||||
| Parameter | Estimate | Std. error | p-value | COR |
| Tree species × Site type × Season × Year (ref. Norway spruce × MT × ES × 2023) | ||||
| Norway spruce × MT × ES × 2024 | −0.307 | 0.412 | 0.457 | 0.736 |
| Norway spruce × MT × MS × 2023 | −0.880 | 0.409 | 0.031 | 0.415 |
| Norway spruce × MT × MS × 2024 | −0.279 | 0.433 | 0.519 | 0.756 |
| Norway spruce × MT × LS × 2023 | 1.774 | 0.526 | 0.001 | 5.894 |
| Norway spruce × MT × LS × 2024 | 0.337 | 0.442 | 0.446 | 1.401 |
| Norway spruce × OMT × ES × 2023 | 0.400 | 0.470 | 0.394 | 1.492 |
| Norway spruce × OMT × ES × 2024 | 0.192 | 0.484 | 0.692 | 1.212 |
| Norway spruce × OMT × MS × 2023 | 0.070 | 0.444 | 0.875 | 1.072 |
| Norway spruce × OMT × MS × 2024 | 2.153 | 0.804 | 0.007 | 8.612 |
| Norway spruce × OMT × LS × 2023 | 1.193 | 0.536 | 0.026 | 3.296 |
| Norway spruce × OMT × LS × 2024 | 0.227 | 0.488 | 0.642 | 1.255 |
| Scots pine × VT × ES × 2023 | −1.573 | 0.369 | <0.001 | 0.207 |
| Scots pine × VT × ES × 2024 | −1.320 | 0.388 | 0.001 | 0.267 |
| Scots pine × VT × LS × 2023 | −0.124 | 0.375 | 0.741 | 0.883 |
| Scots pine × VT × LS × 2024 | −0.622 | 0.374 | 0.096 | 0.537 |
| Seedling anglea (ref. Upright) | ||||
| Askew | −0.940 | 0.125 | <0.001 | 0.391 |
| Prone | −1.376 | 0.292 | <0.001 | 0.253 |
| Soil compactionb (ref. Compacted) | ||||
| Uncompacted | −0.749 | 0.182 | <0.001 | 0.473 |
| Notes (ref. No notes) | ||||
| Tree-top browsingc | −2.296 | 0.298 | <0.001 | 0.101 |
| Waterd | −0.839 | 0.283 | 0.003 | 0.432 |
| Threshold coefficients | ||||
| Dead | Weakened | −4.388 | 0.347 | ||
| Weakened | Healthy | −2.086 | 0.326 | ||
| Random effects | Variance | Std. dev | ||
| Sample plot (n = 60) | 0.147 | 0.384 | ||
| a Upright (<20°), askew (20°–40°), or prone (>40°) b Soil compaction around the planted seedling c Seedlings with eaten tops d The presence of water within the planting spot | ||||
3.4 Seedling height
Norway spruce seedlings planted in early summer and midsummer were the tallest (Fig. 6). In 2023, Norway spruce seedlings planted in midsummer were taller than those planted in early summer. After the second growing season, the difference between planting seasons disappeared. In 2024, healthy Norway spruce seedlings were slightly taller than weakened ones (33.2 ± 6.8 vs. 30.7 ± 6.0 cm). Scots pine seedlings planted in late summer were, on average, taller than those planted in early summer (Fig. 6). Nevertheless, the late summer plantings of Norway spruce and Scots pine are not comparable with those of other planting seasons because of varying seedling sources and initial heights.
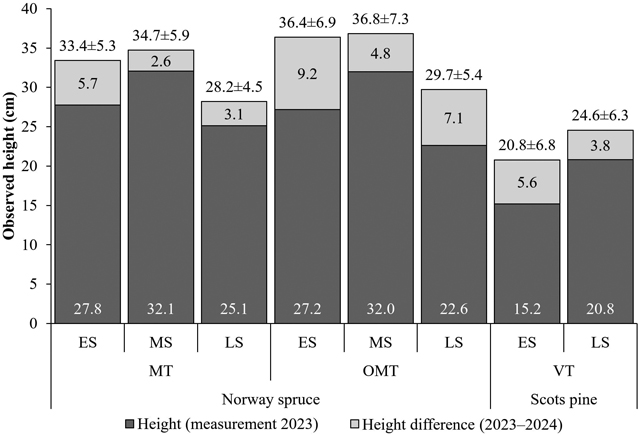
Fig. 6. Observed seedling heights, excluding those that were dead, had broken or eaten tops, or were loose from the soil (2023: n = 1209; 2024: n = 690). The value at the bottom of the dark-gray bar is the mean height in 2023, the value in the light-gray bar is the mean height difference between 2023 and 2024, and the value above the bar is the mean ± standard deviation of height in 2024. Site type: herb-rich heath (Oxalis-Myrtillus type [OMT]), mesic heath (Vaccinium myrtillus type [MT]), and sub-xeric heath (Vaccinium vitis-idaea type [VT]) forests. Planting season: early (ES), mid (MS), and late (LS) summer.
According to the LMEM, site type and the site type × planting season interaction term were not significant predictors of the height of Norway spruce seedlings (Table 6). The planting season alone was a significant predictor. In addition, the height of Norway spruce seedlings was significantly lower on MT than on OMT sites in 2024 (p < 0.05). Seedling vitality (healthy > weakened) was a significant seedling-level predictor (Table 6).
| Table 6. F-tests for the fixed main effects and two-way interaction effects of year, season, and site type in a linear mixed model for Norway spruce height. Year: 2023 and 2024. Site type: herb-rich heath (Oxalis-Myrtillus type [OMT]) and mesic heath (Vaccinium myrtillus type [MT]) forests. Planting season: early (ES), mid (MS), and late (LS) summer. Seedling vitality: healthy and weakened. Degrees of freedom: numerator (numDF) and denominator (denDF). | ||||
| Parameter | numDF | denDF | F-value | p-value |
| Intercept | 1 | 1302 | 790.423 | <0.001 |
| Seedling vitality | 1 | 1302 | 58.274 | <0.001 |
| Year | 1 | 1302 | 97.124 | <0.001 |
| Season | 2 | 30 | 26.932 | <0.001 |
| Site type | 1 | 30 | 0.471 | 0.498 |
| Interaction | ||||
| Year × Season | 2 | 1302 | 16.552 | <0.001 |
| Year × Site type | 1 | 1302 | 39.953 | <0.001 |
| Season × Site type | 2 | 30 | 0.439 | 0.634 |
4 Discussion
4.1 Planting spot quality
Drill-based MSP was performed at three different site types in mineral soil forests throughout the growing season. According to our evaluation, 95% of the prepared planting spots were suitable for planting conifer seedlings, with 61% classified as good and 34% as satisfactory. Consequently, the number of acceptable planting spots was approximately 1710 ha−1, with a target density of 1800 ha−1. This result is in line with that reported by Rantala et al. (2010) for excavator-based mounding (1690 acceptable planting spots ha−1). According to the CLM, the presence of water and ground obstacles, such as logging residues and stones, within the planting spots decreased the odds of good planting spot quality (Table 4). In general, these same factors, especially stones, logging residues, and stumps, reduced the quality of MSP when using other machines (Hämäläinen and Kaila 1987; Andersson et al. 1990; Rantala et al. 2010; Saksa et al. 2018).
Drill-based MSP exposes the soil surface significantly less than conventional MSP methods. For example, the mean area of the planting spots was approximately 80% smaller with drill-based MSP (0.18 m2) than with spot mounding (0.84 m2) and inverting (0.90 m2) (Laine et al. 2020). While drill-based MSP reduces soil exposure, the planting spots need to be fully covered with a layer of pure mineral soil to prevent pine weevil damage (Örlander et al. 1990; Luoranen et al. 2017). Unfortunately, drill-based MSP was unable to fulfill this requirement in all circumstances, as about 13% of the planting spots contained humus layer residues (Table 2). Since humus-rich mounds have been found to increase seedling mortality (Luoranen et al. 2011), this issue should be considered in further development of the drill. Moreover, planting seedlings in sandy pits may by itself increase the risk of pine weevil feeding compared to mounds (Nordlander et al. 2005). Elevated planting spots relative to the ground surface are also preferred at fertile sites because of competitive ground vegetation (Örlander et al. 1996; Luoranen et al. 2007). Therefore, these advantages cannot be achieved with drill-based MSP. Consequently, our findings indicate that drill-based MSP should be primarily used at drier mineral soil sites (e.g., sub-xeric heath forests) where the humus layer is thin.
4.2 Seedling vitality and height
The number of healthy seedlings varied among tree species, planting seasons, site types, and study sites. One year after planting, the mean number of seedlings was higher for healthy Norway spruce seedlings (1794 seedlings ha−1) but lower for Scots pine seedlings (1313 seedlings ha−1) planted in spots prepared by drill-based MSP, compared to those reported by Pikkarainen et al. 2020 for spots prepared using conventional MSP methods (e.g., 1581–1707 Norway spruce seedlings ha−1 and 1686–1731 Scots pine seedlings ha−1). Two years after planting, the mean number of healthy seedlings decreased to 1478 seedlings ha−1 for Norway spruce and to 1054 seedlings ha−1 for Scots pine. Only the Norway spruce seedlings planted in midsummer (OMT) and late summer (MT) were generally successful in 2024 (Fig. 5). As a comparison, Kankaanhuhta et al. (2009) reported that the mean number of Norway spruce seedlings was 1388 seedlings ha−1 three years after planting following patching, mounding, or disc trenching.
Although the target planting density was not achieved at all study sites (e.g., site 3), in some cases, it was significantly exceeded (e.g., site 1). A low planting density can reduce the success of forest regeneration due to natural seedling mortality. Seedling mortality was slightly higher in our study than in previous studies. For example, Sikström et al. (2020) reported survival rates of up to 80%–90% for conifer seedlings planted in conventional MSP spots. In our study, the proportion of healthy seedlings was 59%–82% for Norway spruce and 41%–54% for Scots pine, depending on the site type and planting season (Fig. 4). These findings are consistent with studies that have found Norway spruce plantings to be more successful than Scots pine plantings (Kinnunen 1977; Pikkarainen et al. 2020; Ara et al. 2022; Berglund et al. 2024). Unexpectedly, survival was lowest for Scots pine seedlings planted in early summer, despite this season being highly recommended for planting over late summer due to worse outcomes (Luoranen et al. 2018; Pikkarainen et al. 2020).
Kankaanhuhta et al. (2009) found that the number of planted seedlings was lower on fine-textured mineral soils and peatlands than on medium-textured mineral soils. In fine-textured mineral soils, frost heaving can adversely affect planted seedlings (Örlander et al. 1990; Chantal et al. 2006), particularly when they are planted in late summer (Luoranen et al. 2018). Heiskanen et al. (2013) reported that frost heaving was more common after inverting and ditch mounding than after spot mounding. Thus, in addition to soil properties, how soil is prepared may affect the risk of frost heaving and thereby the survival of planted seedlings. In our study, the mineral soil was classified as medium-textured at all sites, and a few loose seedlings were observed in the follow-up measurements (Table 2). As Nordin et al. (2023) found that the depth-to-water (DTW) index helped to explain the mortality of Scots pine and Norway spruce seedlings planted in mounds, the effect of the DTW index on seedling vitality was also examined in our study. However, the DTW index revealed only a few differences between study sites. Overall, suitable soil properties may at least partly explain the success of seedlings planted in late summer.
Another factor that may have affected seedling vitality is the hot summer weather in 2022 and 2023 (Nordin et al. 2022; Luoranen et al. 2023). The planted seedlings may have experienced drought during the summer, whereas the weather conditions in the fall may have been more favorable for their growth. In addition, differences in seedling production and maintenance (e.g., seedling watering before planting) may have affected the condition of the planted seedlings.
Height of Norway spruce seedlings was associated with seedling vitality, site type, and planting season. Norway spruce seedlings planted in midsummer were taller than those planted in early summer in 2023, most likely because their growing period in the nursery was longer. Nevertheless, this difference diminished by the final follow-up measurement in 2024 (Fig. 6). Healthy seedlings were also taller than weakened ones, as expected. However, analyzing seedling vitality and height development was challenging, as discussed in more detail below.
4.3 Limitations and future direction
Our study considered the most common site types for Norway spruce and Scots pine plantings in Nordic boreal forests. However, it had some limitations in study design, sampling, and dataset. First, the study sites were located close to each other (Fig. 1), resulting in relatively similar site and operating conditions. Therefore, further research is warranted on how different soil properties, such as soil texture and moisture, affect the quality of drill-based MSP and seedling survival. Furthermore, our study did not include control sites in which Norway spruce and Scots pine seedlings were planted in unprepared soil or conventionally prepared soil, such as mounds. Therefore, future research should include such sites to better compare drill-based and conventional MSP methods. Additionally, the original study design for Scots pine late summer plantings had to be changed during the study. Initially, all Scots pine seedlings were to be planted at sites 3 and 4. However, the areas surrounding these sites had already been prepared using other MSP methods before late summer. Thus, a new study site (site 5) was established, where all late summer plantings of Scots pine were eventually conducted.
The planting spots were difficult to distinguish two years after planting, especially when seedlings were weakened or dead. Therefore, not all planted seedlings were found for the final follow-up measurements, which introduced problems into the analysis, as the proportion of healthy seedlings appeared to increase between the first (2023) and second (2024) follow-up measurements. This illogical outcome could have been avoided by marking all measured seedlings and planting spots in the sample plot during the first measurement. Also, variations in seedling sources made comparisons of seedling heights challenging, as initial seedling heights varied with delivery time and age. Moreover, seedlings had different growth periods in the field before the final follow-up measurement (Table 1). Given these limitations, further research should investigate seedling survival and growth in more detail following drill-based MSP.
Finally, the drill’s structure and durability introduced some issues. During the test period in 2022, the drill was slightly damaged by ground obstacles, such as stones, which changed its shape, increasing the diameter and area of the planting spots as the study progressed (Table 3). The machine operator also found the drill to be more difficult to use than conventional excavator-based MSP devices. For example, MSP progress was occasionally hindered by soil and organic matter that stuck on the drill. Thus, the drill design needs to be updated, and its productivity needs to be evaluated accordingly.
Overall, our findings suggest that drill-based MSP is likely most feasible in drier regeneration areas where excessive soil exposure should be avoided, such as in northern boreal regions of Nordic countries where reindeer husbandry is practiced alongside continuous cover forestry (Ersson et al. 2023; Jylhä et al. 2024; Laitila et al. 2025). Moreover, the drill could be tested with different setups and base machines to identify the optimal and most effective concepts. One possible approach would be to use forwarders equipped with the drill in gap-cuttings after logging (Hallikainen et al. 2019) to improve the cost-effectiveness of operations and the multi-use of forestry machinery (Kärhä and Peltola 2004; Ersson et al. 2025).
5 Conclusions
The quality of planting spots prepared using drill-based MSP was sufficient for conifer seedlings. However, ground and soil obstacles, such as roots, stones, stumps, and logging residues, adversely affected planting spot quality. Humus layer residues within planting spots may also increase the risk of pine weevil damage with drill-based MSP. One year after planting, most seedlings were still alive, although the proportion of healthy seedlings decreased notably two years after planting. Nevertheless, uncertainty persisted regarding the vitality and growth of planted seedlings following drill-based MSP. Therefore, future research should test drill-based MSP under varying operating conditions, such as fine-textured mineral soils, and compare its productivity and costs with those of conventional MSP methods, as well as its potential benefits in areas where soil disturbance should be avoided.
Declaration of openness of research materials, data, and code
The data and code are available upon request from Kalle Kemppainen or through the open research repository (https://doi.org/10.5281/zenodo.20917313).
Authors’ contributions
Conceptualization (Eetu Tarvainen, Kalle Kärhä, Markus Strandström), data curation (Eetu Tarvainen, Kalle Kärhä, Kalle Kemppainen, Markus Strandström), formal analysis and investigation (Kalle Kemppainen, Jari Miina), funding acquisition (Kalle Kemppainen, UEF & Luke; Heli Peltola, RCF), methodology (Kalle Kemppainen, Jari Miina), project administration and resources (Ville Kankaanhuhta, Luke; Kalle Kärhä & Heli Peltola, UEF), visualization (Kalle Kemppainen), writing – original draft (Kalle Kemppainen) and writing – review & editing (all).
Declaration of the use of generative artificial intelligence and AI-assisted technologies in the writing process
During the preparation of this article, the corresponding author occasionally used Microsoft Copilot to revise spelling and grammar. Afterward, the corresponding author reviewed and edited the content as needed and assumes full responsibility for the article’s content. The article underwent final proofreading by Cambridge Proofreading LLC.
Acknowledgements
The authors thank Director Tuomo Moilanen from Ponsse Plc for his valuable support for this study. They also thank Heikki Savolainen, Director of Silviculture Operations at Metsähallitus Metsätalous Ltd., for providing the forest regeneration sites used in this study.
Competing interests
The authors declare no competing interests related to this work.
Funding
This study was funded by the Research Council of Finland (formerly the Academy of Finland) through the UNITE Flagship (decision numbers 357906 and 359172). It was also supported by the Faculty of Science, Forestry, and Technology at the University of Eastern Finland and the Natural Resources Institute Finland through 50% salary positions for the doctoral researcher Kalle Kemppainen (from January 2024) under the LUMETO Doctoral Program.
References
Agresti A (2010) Analysis of ordinal categorical data, 2nd ed. John Wiley and Sons, Hoboken, New Jersey. https://doi.org/10.1002/9780470594001.
Äijälä O, Koistinen A, Sved J, Vanhatalo K, Väisänen P (2019) Metsänhoidon suositukset – metsänhoito. [Best practices for sustainable forest management in Finland]. Tapion julkaisuja. https://tapio.fi/projektit/metsanhoidon-suositukset/metsanhoidon-suositusten-aineistopankki/. Accessed 30 September 2025.
Alcázar J, Woodard PM, Rothwell RL (2002) Soil disturbance and the potential for erosion after mechanical site preparation. North J Appl For 19: 5–13. https://doi.org/10.1093/njaf/19.1.5.
Andersson G, Brunberg T, Westerling S (1990) Underlag för produktionsnormer för maskinell markberedning. [Defining productivity standards for mechanized scarification]. The Forest Operations Institute of Sweden, Report 2.
Ara M, Barbeito I, Kalén C, Nilsson U (2022) Regeneration failure of Scots pine changes the species composition of young forests. Scand J For Res 37: 14–22. https://doi.org/10.1080/02827581.2021.2005133.
Berglund M, Öhlund J, Fahlvik N, Johansson F, Ahlinder J, Renman C, Krook M, Vestlund M (2024) Årsrapport. Föryngringskollen Resultat 2023. [Annual report. Regeneration results 2023]. Skogforsk. https://www.skogforsk.se/kunskapsbanken/kunskapsartiklar/2024/foryngringskollen-andra-arets-resultat/. Accessed 30 September 2025.
Björklund N, Nordlander G, Bylund H (2003) Host‐plant acceptance on mineral soil and humus by the pine weevil Hylobius abietis (L.). Agric For Entomol 5: 61–65. https://doi.org/10.1046/j.1461-9563.2003.00163.x.
Cajander AK (1926) The theory of forest types. Acta For Fenn 29, article id 7193. https://doi.org/10.14214/aff.7193.
Chantal M, Rita H, Bergsten U, Löfvenius MO, Grip H (2006) Effect of soil properties and soil disturbance on frost heaving of mineral soil: a laboratory experiment. Can J For Res 36: 2885–2893. https://doi.org/10.1139/x06-181.
Christensen R (2025) Ordinal: regression models for ordinal data. R package version 2025. https://CRAN.R-project.org/package=ordinal. Accessed 17 June 2026.
Christensen R (2026a) Cumulative link models for ordinal regression with the R package ordinal. https://cran.r-project.org/web/packages/ordinal/vignettes/clm_article.pdf. Accessed 27 May 2026.
Christensen R (2026b) A tutorial on fitting cumulative link mixed models with clmm2 from the ordinal package. https://cran.r-project.org/web/packages/ordinal/vignettes/clmm2_tutorial.pdf. Accessed 27 May 2026.
Dumins K, Lazdina D (2018) Forest regeneration quality – factors affecting first year survival of planted trees. In: Proceedings of the 24th Annual International Scientific Conference Research for Rural Development 2018, 16–18 May 2018, Jelgava, Latvia. Latvia University of Life Sciences and Technologies, pp 53–58. https://doi.org/10.22616/rrd.24.2018.008.
Ersson BT, Hansson L, Manner J, Sandström P, Sonesson J (2023) Forest management in northern Fennoscandia: the need for solutions that mitigate conflicts during forest regeneration and increase the use of continuous cover forestry. Silva Fenn 57, article id 23053. https://doi.org/10.14214/sf.23053.
Ersson BT, Manner J, Kärhä K (2025) Forest operations during continuous cover forestry in the Nordics: the need for cost-efficient harvesting solutions like single-machine systems. Silva Fenn 59, article id 25044. https://doi.org/10.14214/sf.25044.
Finnish Meteorological Institute (2024) Weather statistics. https://en.ilmatieteenlaitos.fi/statistics. Accessed 10 March 2024.
Hallikainen V, Hökkä H, Hyppönen M, Rautio P, Valkonen S (2019) Natural regeneration after gap cutting in Scots pine stands in northern Finland. Scand J For Res 34: 115–125. https://doi.org/10.1080/02827581.2018.1557248.
Hämäläinen J, Kaila S (1987) Maaston vaikutus maanmuokkauslaitteiden työjälkeen. [Effect of terrain on work quality of different site preparation machines]. Metsäteho Report 399. https://www.metsateho.fi/maaston-vaikutus-maanmuokkauslaitteiden-tyojalkeen/. Accessed 8 October 2025.
Hansson L, Sten G, Rossander M, Lideskog H, Manner J, van Westendorp R, Li S, Eriksson A, Wallner A, Rönnqvist M, Flisberg P, Edlund B, Möller B, Karlberg M (2024) Autoplant—Autonomous site preparation and tree planting for a sustainable bioeconomy. Forests 15, article id 263. https://doi.org/10.3390/f15020263.
Hechter U, Little KM, Titshall L (2020) The influence of manual and motor-manual pitting implements, pit size and quality on eucalypt performance, South Africa. South For: J For Sci 82: 170–178. https://doi.org/10.2989/20702620.2020.1814109.
Heiskanen J, Rikala R (2006) Root growth and nutrient uptake of Norway spruce container seedlings planted in mounded boreal forest soil. For Ecol Manag 222: 410–417. https://doi.org/10.1016/j.foreco.2005.10.047.
Heiskanen J, Saksa T, Luoranen J (2013) Soil preparation method affects outplanting success of Norway spruce container seedlings on till soils susceptible to frost heave. Silva Fenn 47, article id 893. https://doi.org/10.14214/sf.893.
Heiskanen J, Luoranen J, Viiri H (2019) Maanmuokkauksen vaikutukset taimien kasvuympäristöön. [Effects of site preparation on the seedling growth environment]. In: Laine T, Luoranen J, Ilvesniemi H (eds) Metsämaan muokkaus: Kirjallisuuskatsaus maanmuokkauksen vaikutuksista metsänuudistamiseen, vesistöihin sekä ekologiseen ja sosiaaliseen kestävyyteen. [Forest soil preparation: A literature review on the effects of soil preparation on forest regeneration, water systems, and ecological and social sustainability]. Natural Resource Institute Finland, Natural Resources and Bioeconomy Research 58: 15–24. http://urn.fi/URN:ISBN:978-952-326-813-5.
Hjelm K, Nilsson U, Johansson U, Nordin P (2019) Effects of mechanical site preparation and slash removal on long-term productivity of conifer plantations in Sweden. Can J For Res 49: 1311–1319. https://doi.org/10.1139/cjfr-2019-0081.
Jylhä P, Laitila J, Hallikainen V, Aatsinki P, Kukkonen M, Hilli A, Karvonen L, Rautio P (2024) Seedling damage caused by wood harvesting and soil scarification in rotation and continuous cover forestry in Scots-pine-dominated boreal forests. Scand J For Res 39: 44–58. https://doi.org/10.1080/02827581.2023.2289656.
Kankaanhuhta V, Saksa T, Smolander H (2009) Variation in the results of Norway spruce planting and Scots pine direct seeding in privately-owned forests in southern Finland. Silva Fenn 43: 51–70. https://doi.org/10.14214/sf.217.
Kärhä K, Peltola J (2004) Metsäkoneiden monikäyttöisyys. [Versatility of forest machines]. Metsäteho Report 181. https://www.metsateho.fi/metsakoneiden-monikayttoisyys/. Accessed 17 June 2026.
Karjalainen E (2006) The visual preferences for forest regeneration and field afforestation – four case studies in Finland. Diss For 31. https://doi.org/10.14214/df.31.
Kinnunen K (1977) Istutuksen onnistuminen ja taimistojen alkukehitys Länsi-Suomen yksityismetsissä. [The survival and initial development of plants in private forests in Western Finland]. Folia For 318: 1–34. http://urn.fi/URN:ISBN:951-40-0287-3.
Laine T, Kankaanhuhta V, Rantala J, Saksa T (2020) Effects of spot mounding and inverting on growth of conifers, exposed mineral soil and natural birch regeneration. Silva Fenn 54, article id 10369. https://doi.org/10.14214/sf.10369.
Laitila J, Repola J, Holmström E (2025) Time consumption models for predicting harvester productivity when selection cutting, thinning from below, and clearcutting Scots pine-dominated stands in Finnish Lapland. Int J For Eng 36: 178–193. https://doi.org/10.1080/14942119.2024.2448935.
Lidman FD, Karlsson M, Lundmark T, Sängstuvall L, Holmström E (2024) Birch establishes anywhere! So, what is there to know about natural regeneration and direct seeding of birch? New For 55: 157–171. https://doi.org/10.1007/s11056-023-09971-z.
Löf M, Dey DC, Navarro RM, Jacobs DF (2012) Mechanical site preparation for forest restoration. New For 43: 825–848. https://doi.org/10.1007/s11056-012-9332-x.
Luoranen J, Saksa T, Finér L, Tamminen P (2007) Metsämaan muokkausopas. [Site preparation manual for forest lands]. Finnish Forest Research Institute (Metla), Suonenjoki. http://urn.fi/URN:ISBN:978-951-40-2059-9.
Luoranen J, Rikala R, Smolander H (2011) Machine planting of Norway spruce by Bracke and Ecoplanter: an evaluation of soil preparation, planting method and seedling performance. Silva Fenn 45: 41–357. https://doi.org/10.14214/sf.107.
Luoranen J, Viiri H, Sianoja M, Poteri M, Lappi J (2017) Predicting pine weevil risk: effects of site, planting spot and seedling level factors on weevil feeding and mortality of Norway spruce seedlings. For Ecol Manage 389: 260–271. https://doi.org/10.1016/j.foreco.2017.01.006.
Luoranen J, Saksa T, Lappi J (2018) Seedling, planting site and weather factors affecting the success of autumn plantings in Norway spruce and Scots pine seedlings. For Ecol Manage 419–420: 79–90. https://doi.org/10.1016/j.foreco.2018.03.040.
Luoranen J, Riikonen J, Saksa T (2021) Factors affecting winter damage and recovery of newly planted Norway spruce seedlings in boreal forests. For Ecol Manage 503, article id 119759. https://doi.org/10.1016/j.foreco.2021.119759.
Luoranen J, Riikonen J, Saksa T (2023) Damage caused by an exceptionally warm and dry early summer on newly planted Norway spruce container seedlings in Nordic boreal forests. For Ecol Manage 528, article id 120649. https://doi.org/10.1016/j.foreco.2022.120649.
McEwan A, Steenkamp J (2014) Silviculture modernization in the South African forestry industry. In: Ciancio O (ed) Proceedings of the Second International Congress of Silviculture, 26–29 November 2014, Florence, Italy. Accademia Italiana di Scienze Forestali, Florence, pp 822–826. https://doi.org/10.4129/2cis-ame-sil.
Mehtätalo L, Lappi J (2020) Biometry for forestry and environmental data: with examples in R, 1st ed. CRC Press, Boca Raton and Oxon. https://doi.org/10.1201/9780429173462.
Natural Resources Institute Finland (2025) Silvicultural and forest improvement work in 2024 – statistics database. https://www.luke.fi/en/statistics/silvicultural-and-forest-improvement-work/silvicultural-and-forest-improvement-work-2024. Accessed 26 September 2025.
Nilsson U, Örlander G (1999) Vegetation management on grass-dominated clearcuts planted with Norway spruce in southern Sweden. Can J For Res 29: 1015–1026. https://doi.org/10.1139/x99-071.
Nordin P, Olofsson E, Hjelm K (2022) Successful spruce regenerations – impact of site preparation and the use of variables from digital elevation models in decision-making? Scand J For Res 37: 33–44. https://doi.org/10.1080/02827581.2022.2028895.
Nordin P, Olofsson E, Hjelm K (2023) Within-site adaptation: growth and mortality of Norway spruce, Scots pine and silver birch seedlings in different planting positions across a soil moisture gradient. Silva Fenn 57, article id 23004. https://doi.org/10.14214/sf.23004.
Nordlander G, Bylund H, Björklund N (2005) Soil type and microtopography influencing feeding above and below ground by the pine weevil Hylobius abietis. Agric For Entomol 7: 107–113. https://doi.org/10.1111/j.1461-9555.2005.00257.x.
Örlander G, Gemmel P, Hunt J (1990) Site preparation: a Swedish overview. FRDA Report 105, British Columbia, Canada. ISBN 0-7726-1081-9.
Örlander G, Nilsson U, Hällgren J-E (1996) Competition for water and nutrients between ground vegetation and planted Picea abies. New Zeal J For 26: 99–117.
Örlander G, Hallsby G, Gemmel P, Wilhelmsson C (1998) Inverting improves establishment of Pinus contorta and Picea abies – 10-year results from a site preparation trial in northern Sweden. Scand J For Res 13: 160–168. https://doi.org/10.1080/02827589809382972.
Palviainen M, Finér L, Laurén A, Launiainen S, Piirainen S, Mattsson T, Starr M (2014) Nitrogen, phosphorus, carbon, and suspended solids loads from forest clear-cutting and site preparation: long-term paired catchment studies from eastern Finland. Ambio 43: 218–233. https://doi.org/10.1007/s13280-013-0439-x.
Piirainen S, Finér L, Mannerkoski H, Starr M (2009) Leaching of cations and sulphate after mechanical site preparation at a boreal forest clear-cut area. Geoderma 149: 386–392. https://doi.org/10.1016/j.geoderma.2009.01.003.
Pikkarainen L, Luoranen J, Kilpeläinen A, Oijala T, Peltola H (2020) Comparison of planting success in one-year-old spring, summer and autumn plantings of Norway spruce and Scots pine under boreal conditions. Silva Fenn 54, article id 10243. https://doi.org/10.14214/sf.10243.
Pinheiro J, Bates D (2026) Nlme: linear and nonlinear mixed effects models. R package version 3.1-169. https://cran.r-project.org/package=nlme. Accessed 17 June 2026.
R Core Team (2025) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.r-project.org/. Accessed 28 July 2025.
Ramantswana M, Guerra SPS, Ersson BT (2020) Advances in the mechanization of regenerating plantation forests: a review. Curr For Rep 6: 143–158. https://doi.org/10.1007/s40725-020-00114-7.
Rantala J, Saarinen VM, Hallongren H (2010) Quality, productivity and costs of spot mounding after slash and stump removal. Scand J For Res 25: 507–514. https://doi.org/10.1080/02827581.2010.522591.
Ring E, Sikström U (2024) Environmental impact of mechanical site preparation on mineral soils in Sweden and Finland – a review. Silva Fenn 58, article id 23056. https://doi.org/10.14214/sf.23056.
Saksa T, Miina J, Haatainen H, Kärkkäinen K (2018) Quality of spot mounding performed by continuously advancing mounders. Silva Fenn 52, article id 9933. https://doi.org/10.14214/sf.9933.
Sikström U, Hjelm K, Holt Hanssen K, Saksa T, Wallertz K (2020) Influence of mechanical site preparation on regeneration success of planted conifers in clearcuts in Fennoscandia – a review. Silva Fenn 54, article id 10172. https://doi.org/10.14214/sf.10172.
Skogsstyrelsen (2025) Silvicultural activities in 2024. https://www.skogsstyrelsen.se/en/statistics/silvicultural-activities/silvicultural-activities/. Accessed 26 September 2025.
Smolander A, Heiskanen J (2007) Soil N and C transformations in two forest clear-cuts during three years after mounding and inverting. Can J Soil Sci 87: 251–258. https://doi.org/10.4141/S06-028.
Sutton RF (1993) Mounding site preparation: a review of European and North American experience. New For 7: 151–192. https://doi.org/10.1007/BF00034198.
Thiffault N, Nordin P, Wotherspoon A, Hjelm K, Olofsson E (2025) A trans-Atlantic perspective on successful plantation establishment in boreal ecosystems: lessons learned and research opportunities. New For 56, article id 16. https://doi.org/10.1007/s11056-024-10086-2.
Uotila K, Rantala J, Saksa T, Harstela P (2010) Effect of soil preparation method on economic result of Norway spruce regeneration chain. Silva Fenn 44: 511–524. https://doi.org/10.14214/sf.146.
Uotila K, Luoranen J, Saksa T, Laine T, Heiskanen J (2022) Long-term growth response of Norway spruce in different mounding and vegetation control treatments on fine-textured soils. Silva Fenn 56, article id 10762. https://doi.org/10.14214/sf.10762.
Zulu TB, Ramantswana MM, Spinelli R (2024) Mechanical site preparation in South Africa: comparing the productivity of pitting machine operators under different site conditions. Int J For Eng 35: 362–370. https://doi.org/10.1080/14942119.2024.2373011.
Total of 66 references.
