Jaana Luoranen  ,
Heli Viiri
,
Heli Viiri
Comparison of the planting success and risks of pine weevil damage on mineral soil and drained peatland sites three years after planting
Luoranen J., Viiri H. (2021). Comparison of the planting success and risks of pine weevil damage on mineral soil and drained peatland sites three years after planting. Silva Fennica vol. 55 no. 4 article id 10528. https://doi.org/10.14214/sf.10528
Highlights
- The planting success was poorer on peatland sites than on mineral soil
- Dense ground vegetation cover is more probable on peatland than on mineral soil
- No differences in pine weevil feeding damage on mineral soil and peatland were found
- Cultivated mineral soil reduced the vegetation cover, feeding damage and seedling mortality.
Abstract
Over 20% of regeneration operations will be on drained peatland in the next decade in Finland. There are only a few studies comparing the planting success and the risk of pine weevil (Hylobius abetis (L.) feeding damage on mineral soil and drained peatland. Thirty sites planted with Norway spruce (Picea abies (L.) H. Karst.) container seedlings in 2009 in Southern and Central Finland were inventoried three growing seasons after planting. Prediction models for the probability of survival, pine weevil damage and the presence of ground vegetation cover were done separately for peatland and mineral soil sites. The planting success was 17% lower on peatland sites (1379 surviving seedlings ha–1) than on mineral soil (1654 seedlings ha–1). The factors explaining the survival were the ground vegetation cover and type of the planting spot on the peatland, and the ground vegetation cover on mineral soil. On mineral soil, 76% of the planting spots were on cultivated mineral soil while on peatland only 28% of the seedlings were planted on similar spots. There were also fewer seedlings that were surrounded by dense ground vegetation on mineral soil (4%) than on peatland (14%). Pine weevil feeding damage did not differ significantly on peatland (23%) or mineral soil (18%). The more time there was from clear-cutting, the more the probability of pine weevil feeding damage was reduced on both soil classes. Additionally, cover vegetation in the vicinity of the seedlings increased on mineral soil. Cultivated planting spots, especially those covered by mineral soil, prevented pine weevil feeding and reduced the harmful effects of vegetation on the seedlings both on mineral soil and peatland.
Keywords
survival;
planting;
mechanical site preparation;
ground vegetation;
forest regeneration;
Hylobius abietis
-
Luoranen,
Natural Recourses Institute Finland (Luke), Production systems, Juntintie 154, FI-77600 Suonenjoki, Finland
 https://orcid.org/0000-0002-6970-2030
E-mail
jaana.luoranen@luke.fi
https://orcid.org/0000-0002-6970-2030
E-mail
jaana.luoranen@luke.fi
-
Viiri,
UPM Forest, Peltokatu 26 C, P.O. Box 85, FI-33100 Tampere, Finland
https://orcid.org/0000-0003-3952-9481
E-mail
heli.viiri@upm.com
Received 19 February 2021 Accepted 23 August 2021 Published 25 August 2021
Views 60270
Available at https://doi.org/10.14214/sf.10528 | Download PDF

1 Introduction
Peatlands cover over 20% of the land area of Europe and 24% of European peatlands are in the Baltic Sea basin, especially in Estonia, Finland, and Sweden (Vasander et al. 2003). According to Vasander et al. (2003), the country with the highest proportion of peatlands in the world is Finland, which has over 8 Mha of forested peatlands, of which 4.65 Mha is drained (Korhonen et al. 2017). During the next decade, 22% of regeneration operations will take place on drained peatlands (Korhonen et al. 2017). Continuously covered forestry (CCF) is largely considered as an alternative to even-aged management with clear-cut, mechanical site preparation and planting or seeding in Nordic boreal drained peatlands (Nieminen et al. 2018). CCF on drained peatlands is still being studied, but based on the literature, Nieminen et al. (2018) consider that it could be an economically and environmentally feasible management option. With CCF, it might be possible to reduce greenhouse gas emissions and negative water quality impacts (Nieminen et al. 2018). However, there are not good enough conditions for the successful natural regeneration needed for CCF on all drained peatlands. This is especially the case for older drained peatlands with a thin peat layer and when Sphagnum mosses have disappeared (Saarinen 2013) and when there is root-rot infection in the stand (Silver and Piri 2017). Peatlands and their regeneration success are important for forestry in the Baltic Sea basin. Artificial regeneration, including planting, is a potential regeneration method which could be applied also in the future.
For Nordic boreal forests, comparisons of the differences in the planting success of mineral soil and peatland sites are rare. Kankaanhuhta et al. (2009) studied the results of Norway spruce (Picea abies (L.) H. Karst.) planting in privately-owned forests in Southern Finland. In their study, the number of surviving seedlings was 10% lower when the soil class was peat rather than medium coarse mineral soil three years after planting.
In boreal forests, one of the most important agents that reduces the planting success of conifer seedlings are large pine weevils (Hylobius abietis L.) (Långström 1982; Nordlander et al. 2011). In addition to leading to increased mortality, pine weevil damage can also impair the growth of seedlings (Luoranen et al. 2017). The surrounding soil texture affects the risk of pine weevil feeding. Exposed mineral soil around the seedling strongly reduces the probability of pine weevil feeding (Björklund et al. 2003), and this also applies to drained peatlands with a thin peat layer (Luoranen and Viiri 2012). The damage caused by pine weevils has largely been studied on mineral soil sites (e.g., Långström 1982; Nordlander et al. 2011; Luoranen et al. 2017). However, large-scale pine weevil damage has also been observed on peatlands (Hånell 1993; Holgén and Hånell 2000; Pearson et al. 2011; Hytönen et al. 2020), but there are few studies comparing pine weevil damage on peatland and mineral soil sites (Luoranen and Viiri 2012). Luoranen and Viiri (2012) showed that pine weevil damage was marginally more probable on peatland than mineral soil sites one growing season after planting. In Southern Finland, conifer seedlings are protected against pine weevils with insecticides in the nursery before planting. The use of insecticides is prohibited in the forest and other protection methods such as sand-coating are also not used. The protection effect of insecticides is reduced after the first growing season (Viiri et al. 2007) and pine weevil feeding can continue for several years after clear-cutting and planting in boreal forests (Långström 1982; Piri et al. 2020). Thus, survey results after the first growing season may underestimate the pine weevil feeding damage.
The ground vegetation cover increases with time after a clear-cut (Nilsson and Örlander 1995; Örlander et al. 1996), especially on fertile peatlands (Moilanen et al. 1995). Herbaceous vegetation and deciduous tree seedlings compete with planted seedlings for water, nutrients, and light, reducing growth or increasing seedling mortality (Nilsson and Örlander 1995; Örlander et al. 1996). In addition, high vegetation cover increases pine weevil feeding (Örlander and Nordlander 2003; Petersson et al. 2006) and vole damage (Teivainen et al. 1986; Huitu et al. 2013). One of the most important effects of site preparation is to reduce the competition of ground vegetation (Nilsson and Örlander 1995; Örlander et al. 1996).
To support decisions on various operations and their timing, such as soil preparation in the regeneration chain, it is important to understand the impacts of various factors on pine weevil feeding damage and the success of regeneration, as well as the differences in these impacts on peat and mineral soil sites. The aim of the current study was to compare the planting success and the risk of pine weevil feeding damage on peatland and mineral soil sites three years after planting. We also assessed several factors related to the regeneration chain and seedling microsites. Prediction models were fitted to the data aiming to clarify factors which would predict the survival of the seedlings as well as the pine weevil feeding damage and the ground vegetation cover on peatland and mineral soil.
2 Materials and methods
2.1 Study sites and sampling
A study by Luoranen and Viiri (2012) included 60 regeneration sites, one half was planted in 2008 and the other half in 2009. Thirty sites planted in 2009 were selected for the present study. These sites are located in the Pirkanmaa region (regions A and B) and in the South Savo region (region C) of Southern and Central Finland, ranging between latitudes 61 and 63°N (Fig. 1). There were 0–3 years between clear-cutting and planting and the age of the clear-cuts at time of the inventory varied from 3 to 6 years (Table 1). Norway spruce container seedlings were planted in spring 2009. All the seedlings were treated with insecticides (active ingredient: deltamethrin or lambda-syhalotrin) against pine weevil feeding before planting. All the peatland sites had been drained and were either transformed peatland or being transformed with a very thin peat layer (Luoranen and Viiri 2012). Based on the seedling appearance, about 1% of the seedlings were presumed to have been supplementarily planted in regions A and B.
Fig. 1. The location of the survey sites in Finland. Triangles are for mineral soil and circles for peatland sites. Regions A, B and C were operated by different organizations.
| Table 1. Numbers of regeneration sites and inventoried planting spots in peatland and mineral soils (peatlands are presented first) included in the survey done three years after planting in Central Finland. The proportion of seedlings (percentage of seedlings belonging to each category) checked in each geographical area in each category of sample plot-level variables are also presented. | |||
| Geographical area | |||
| Region Aa | Region B | Region C | |
| The number of regeneration sites | 4/6 | 5/5 | 7/3 |
| The number of inventoried planting spots | 369/612 | 474/448 | 611/267 |
| Number of inventoried planting spots ha–1 | 1845/1943 | 1896/1792 | 1721/1780 |
| Site typeb, % of planting spots | |||
| Mesic (Oxalis-Myrtillus type) | 14/0 | ||
| Sub-mesic (Myrtillus type) | 51/81 | 60/95 | 75/84 |
| Sub-dry (Vaccinium type) | 49/19 | 40/5 | 11/16 |
| Soil texture type, % of planting spots | |||
| Medium coarse mineral soil | 14/65 | 3/83 | 14/67 |
| Fine mineral soil | 3/17 | 0/17 | 1/0 |
| Peat | 83/18 | 97/0 | 85/33 |
| Stoniness, % of planting spots | |||
| No stones | 90/77 | 96/51 | 86/69 |
| Normal stoniness | 10/22 | 4/34 | 3/16 |
| Very stony | 0/1 | 0/15 | 5/15 |
| Site preparation method, % of planting spots | |||
| Soil inversion | 83/100 | 13/55 | |
| Spot mounding | 34/86 | 1/0 | 25/12 |
| Ditching and mounding | 66/14 | 16/0 | 62/33 |
| Clear-cut age, % of planting spots | |||
| 3 years | 21/21 | ||
| 4 years | 58/56 | 71/31 | |
| 5 years | 47/67 | 21/23 | 29/69 |
| 6 years | 53/33 | ||
| a Geographical regions A and B were in the Pirkanmaa and region C in South Savo in Finland. b Site type classification is based on Tonteri et al. 1990 and Cajander 1949. | |||
Systematic plot sampling was executed as in Luoranen and Viiri (2012) but the location of the sample plots on the sites were not the same as in the first inventory. The sampling was based on an area of the regeneration site which varied between 0.5 ha and 6 ha. On average, ten circular sample plots (50 m2, radius 3.99 m) were systematically placed per site. The distance between sampled plots was shorter on the smaller sites than on the larger sites. The same person carried out the inventory on all sites in regions B and C in October 2011 and in region A in May 2012.
On each plot, the site type (mesic, sub-mesic and sub-dry based on Tonteri et al. 1990), soil texture type and stoniness were visually determined (Table 1). According to the World Reference Base, the soil type in this study was mainly podzol and the soil texture was assessed visually and classified into four categories based on the main soil particle size in mineral soil and the amount of organic material. The four categories were: coarse mineral soil (grain size is easy to evaluate by naked eye), medium coarse mineral soil (single grains can be still detect by naked eye, grains detached), fine mineral soil (single soil grains cannot be detect by naked eye), and peat. In Luoranen and Viiri (2012), a site was classified as a peatland soil class category when most of the soil texture observations on the site were peat, otherwise the site was classified as a mineral soil. We used the same categories here for each site. The sites were also classified according to the visible stoniness into three categories. The classifications were: no stones, normal stoniness or very stony. For each seedling within a plot, the mechanical site preparation (MSP) method, type of planting spot, the seedling condition (healthy, slightly damaged, weakened, dead) and the cause (pine weevil, vole, other reason) of damage, as well as the ground vegetation cover were recorded. If pine weevil feeding damage was minor without affecting further development of the seedling, the seedling’s condition was classified as healthy. We used four categories for the MSP: patching (humus layer removed), soil inversion (humus and mineral soil layers were turned over on the same spot from which they were taken, with mineral soil on top of inverted humus), spot mounding (humus and underlying mineral soil turned over onto the adjacent undisturbed forest floor, with the mound consisting of a double humus layer) and ditch mounding (material for the mounds was taken from excavated ditches and dropped onto the undisturbed humus). There were five categories for the type of planting spot (spots in which a seedling had been planted): undisturbed peat, undisturbed mineral soil, cultivated peat (including patches and mounds covered with peat), cultivated humus (mounds covered with humus) or cultivated mineral soil (patches and mounds covered with mineral soil). The ground vegetation cover was assessed visually and included the cover vegetation, herbaceous plants, and naturally born tree seedlings that grew in a half meter radius around the planted seedling. Based on potential effects on the planted seedling, ground vegetation cover was classified into categories: no vegetation, sparse vegetation (some plants without any effect on the seedling), and dense vegetation (vegetation affecting seedling development). The age of the clear-cut (the years since clear-cutting at the time of the inventory) was used as background information.
In total, 2781 seedlings in three regions at 30 sites were evaluated (Table 1). Most of the sites were classified to be sub-mesic, and 48% were on mineral soil and 52% on peatland. On the mineral soil sites, 67% had no stones, 25% had a normal level of stoniness and 8% were very stony. The MSP methods applied were soil inversion, spot mounding, and ditch mounding for 33, 19 and 48%, respectively, of the inventoried seedlings on peatland. The corresponding values were 45, 42 and 13%, respectively, on mineral soil.
The data presented in Luoranen and Viiri (2012) was used to calculate the number of planted seedlings and the surviving seedlings per hectare after the first growing season. The current data was used to calculate the number of planting spots and surviving seedlings per hectare three years after planting. A seedling was defined to be surviving if its condition was either healthy or just slightly damaged.
2.2 Statistical analysis
First, we analyzed the difference in seedling survival (defined as healthy or only slightly damaged seedlings), pine weevil feeding damage, vole damage, and mortality caused by pine weevil feeding between soil class categories, i.e., on peatland and mineral soil. Additionally, the differences in the ground vegetation around a seedling (combined categories for sparse and dense vegetation) were analyzed for the two soil classes. In the generalized linear mixed (GLMM) models, the soil class was used as a fixed effect and the geographical area, regeneration site and sample plot within a site were set as random effects. All GLMM analyses were done using the PROC GLIMMIX software in SAS for Windows 9.4 (SAS Institute Inc., Cary, NC, USA).
The prediction models were used to calculate the probability of survival (healthy or only slightly damaged seedlings), pine weevil feeding damage, and the presence of ground vegetation three years after planting. All seedlings with any feeding damage (including all seedling conditions classes except healthy ones) were used in the prediction model for pine weevil feeding damage. Supplementary planted seedlings were included but empty planting spots were excluded in the analysis.
We used the same approach as previously used in Luoranen et al. (2018) and the model structure is described there. The analysis was conducted with binary data using the Laplace approximation method and the logit link function. The regeneration site and sample plot within a regeneration site were used as random effects. All assessed site-, sample plot- and seedling-level variables were used as potential predictor variables.
In the prediction models, explanatory variables differed between soil classes and separate fixed parameters were modelled for peatland and mineral soil using dummy variables. Additionally, the variances of the random effects were assumed to be different for the soil classes. In the first run, when there were only intercepts in the model, the geographical area was set as a random effect. In all cases standard deviations for that level were so small that we excluded them from further runs. The modelling was done by adding variables (mentioned in Tables 1 and 2) one by one, leaving only the significant ones (p ≤ 0.05) in the final models. The models were compared using fit statistics in the GLIMMIX outputs: smaller values of a -2 Log Likelihood were considered to explain most of the variation. Additionally, many variables correlated and only the most logical ones were kept in the final model. After the first analysis, no differences between classes for several variables were found and the classes were combined keeping the model as simple as possible. An inverse link function was used to transform the linear predictor back to probabilities. Random effects were taken to be zero in the linear predictor.
There were three levels of variables in the final models: (1) the clear-cut age assessed on a regeneration site level, (2) the site type and MSP at the sample plot level, and (3) the type of the planting spot and the cover of ground vegetation separately for each seedling.
The coefficient of determination (CD) was computed as (1-var/var0), where var was the variance of the random effect and the var0 was the variance of the random effect in the model where the intercept was the only fixed effect.
The differences in the number of planting spots or surviving seedlings per hectare between soil classes and geographical areas were analyzed using a univariate analysis of variance in IBM SPSS Statistics, Version 25.
3 Results
3.1 Number of planting spots and surviving seedlings
Most of seedlings were planted in cultivated planting spots (mainly on mounds, some on patches), both on peatland and mineral soil sites (Table 2). On peatland, most of the seedlings were planted in cultivated peat, but 28% were planted in cultivated mineral soil, which is explained by the thin peat layer common among the sites included in this study. On mineral soil, 7% of the seedlings were planted in cultivated humus (mounds covered by humus). Approximately 2% of the seedlings were planted in undisturbed soil both on mineral soil and peatland.
| Table 2. The proportion of Norway spruce seedlings (percentage) belonging to each category of seedling-level variables presented separately for each geographical area on peatland and mineral soils. Peatlands are presented first. In the last column, the average proportions of seedlings belonging to each category on each soil class in the whole data set are presented. Regions are explained in Table 1. | ||||
| Geographical area | ||||
| Region A | Region B | Region C | Average | |
| Number of surviving seedlings ha–1 | 1647/1847 | 1366/1541 | 1237/1440 | 1379/1654 |
| Type of planting spot, % of spots | ||||
| Undisturbed peat | 0.5/0 | 1.5/0 | 3.5/0.5 | 2/0.2 |
| Undisturbed mineral soil | 0/0 | 0/6 | 0.2/1.5 | 0.1/2.5 |
| Cultivated peat | 67/15.5 | 64/0 | 74/30 | 69/13 |
| Cultivated humus | 1.5/5.5 | 0.5/9.5 | 0.2/9 | 0.5/7.5 |
| Cultivated mineral soil | 31/79 | 34/84.5 | 22/58 | 28/76 |
| Ground vegetation cover, % of spots | ||||
| No vegetation | 31/37.5 | 42/42 | 14.5/35 | 29/39 |
| Sparse | 64/61 | 49/53 | 58/60 | 57/57 |
| Dense | 5/1.5 | 9/5 | 27.5/5 | 14/4 |
| Degree of damage, % of seedlings | ||||
| Healthy | 83/90 | 44/56 | 46/46 | 55/69.5 |
| Slightly damaged | 5/4 | 26/28 | 23/33 | 20/18 |
| Weakened | 10/5 | 26/13 | 25/17 | 21/10 |
| Dead | 1/0 | 1/0.5 | 2/0.5 | 1/0.5 |
| Empty mound | 2/1 | 4/2.5 | 4/3.5 | 3/2 |
| Cause of damage, % of seedlings | ||||
| Pine weevil | 6/5 | 39/31 | 22/28 | 23/18 |
| Vole | 0/0 | 5/1 | 23/17 | 11/4 |
| Other reasons | 9/4 | 15/11 | 10/7 | 11/7 |
After the first growing season, there were 1852 ± 347 (mean ± standard deviation) planting spots ha–1 and 1839 ± 347 surviving seedlings ha–1 with no differences between soil classes or geographical areas (p values between 0.197 and 0.889). After three years, approximately 1830 ± 331 planting spots ha–1 with no differences between the soil classes (p = 0.705) were found. The number of spots ha–1 was lower in the C region than in the other regions (Table 1; p = 0.003).
The seedling mortality was low in all regions (Table 2) and the probability of death did not differ between the soil classes (p = 0.351). However, there were approximately 21% and 10% weakened seedlings on peatland and mineral soil, respectively, meaning that the survival probability of the seedlings was lower on peatland (with a predicted probability of 0.83) than on mineral soil (0.95; p = 0.007). This also lowered the number of surviving seedlings per ha more on peatland than on mineral soil sites (p = 0.001) in all regions three years after planting (Table 2). The number of surviving seedlings per ha was also lower in regions B and C than in region A (p < 0.001). The number of surviving seedlings per ha within a site varied from 500 to 1800 on peatland, and from 1100 to 1900 on mineral soil (Fig. 2). There were ≥ 1500 surviving seedlings per ha on 44% of the sites on peatland, whereas the corresponding value was 79% for sites on mineral soil.
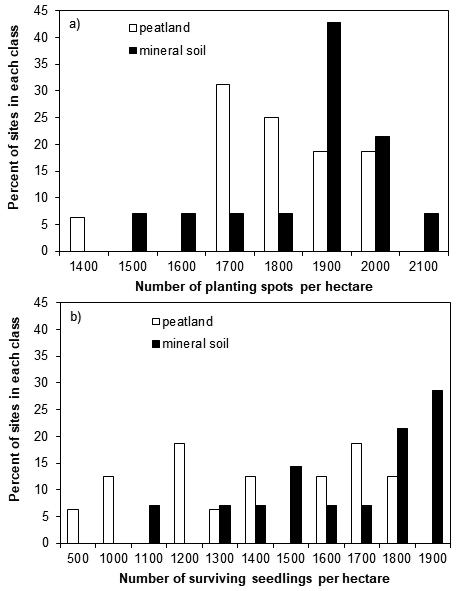
Fig. 2. The number of a) found planting spots and b) surviving Norway spruce seedlings per hectare at regeneration sites on peatland and mineral soil sites three years after planting. Each bar presents the percentage of sites belonging to each class.
3.2 Seedling damage
For most of damaged seedlings, the cause of damage was pine weevil feeding (Table 2). It caused 23 and 18% of all the seedling damage on peatland and mineral soil. The corresponding values for vole damage were 11 and 4%. Both pine weevil and vole damage varied substantially between geographical areas. Other reasons caused on average 9% of observed damage and the cause of these reasons was unknown in most cases. The recognized reasons included frost heaving (1%) and moose trampling (1%). Damaged shoots (multiple or dry shoots) were observed for 2% of the seedlings.
There was a large amount of variation in the vole damage between the regeneration sites (Table 2) and the estimate of the standard deviation at the regeneration site level was 15. When all regions were included, the predicted probability of vole damage was 0.0004 and 0.003 on mineral soil and peatland, respectively, without any statistically significant differences between soil classes (p = 0.206). In region C, the probability of vole damage was higher, and the probabilities were 0.06 and 0.07 for peatland and mineral soil without statistically significant differences between the soil classes (p = 0.686). Vole damage was the main reason for the mortality of seedlings causing 50 and 17% of the mortality on peatland and mineral soil sites without any statistically significant difference (p = 0.491).
3.3 Prediction models
3.3.1 Seedling survival
On peatland, the ground vegetation cover and the type of the planting spot explained the probability of survival (Table 3). The ground vegetation cover predicted survival on mineral soil. The survival probability was lower when a seedling was surrounded by dense vegetation in comparison to other vegetation classes both on peatland and mineral soil (Fig. 3). On peatland, the survival probability was also lower when a seedling had been planted in cultivated peat than in cultivated mineral soil. The variances in geographical level were small, and they were dropped from the final model. The CDs of the model were always less than 0.25 in all levels. The model explained the variation best at the regeneration site-level for peatland (Table 3).
| Table 3. The model for the survival probability of Norway spruce seedlings three years after planting in peatland (Peat) and mineral soil (Mineral) sites. Estimates, their standard errors (SE) and the significance of parameters of fixed effects and the variances of random effects (regeneration site and sample plot within a regeneration site) in a generalized linear mixed model are given. Soil classes Peat and Mineral were used as dummy variables. The fixed and random effect variances were estimated separately for each of them. In the final model, the fixed effects were the type of planting spot (reference category “Cultivated mineral soil”) and the ground vegetation cover around a seedling (reference category “No vegetation”). CD is the coefficient of determination for different random effects. | ||||
| Effect type | Estimate (SE) | t Value | p-value | |
| Fixed effects | ||||
| Peat | 1.87 (0.37) | 5.01 | <0.001 | |
| Peat × Ground vegetation cover | Dense | –0.87 (0.28) | –3.07 | <0.001 |
| Sparse | 0.29 (0.20) | 1.49 | 0.137 | |
| Peat × Type of planting spot | Undisturbed spot | –0.58 (0.53) | –1.09 | 0.276 |
| Cultivated organic spot | –0.61 (0.27) | –2.28 | 0.023 | |
| Mineral | 3.12 (0.47) | 6.69 | <0.001 | |
| Mineral × Ground vegetation cover | Dense | –1.09 (0.47) | –2.31 | 0.021 |
| Sparse | 0.13 (0.25) | 0.53 | 0.597 | |
| Variances of random effects | Estimate (SE) | CD | ||
| Regeneration site (Peat) | 0.99 (0.43) | 0.24 | ||
| Sample plot (Peat) | 0.54 (0.18) | 0.13 | ||
| Regeneration site (Mine) | 1.77 (0.97) | –0.05 | ||
| Sample plot (Mine) | 1.26 (0.42) | 0.13 | ||
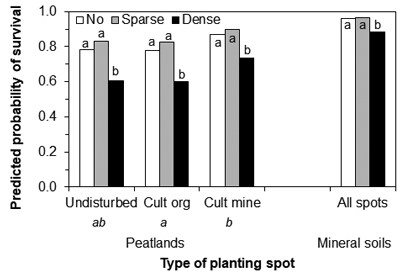
Fig. 3. The predicted probability of Norway spruce seedling survival (healthy and slightly damaged) on the different types of planting spots (only for peatland) and the ground vegetation cover classes on peatland and mineral soil sites. Undisturbed = planting spot on undisturbed soil, Cult org = cultivated (mound or patch) planting spot covered by peat, Cult mine = cultivated (mound or patch) planting spot covered by mineral soil. Letters below the type of planting spot on peatland indicate the statistically significant differences between types. Letters above bars indicate statistically significant differences between ground vegetation classes within a type of planting spot.
3.3.2 Pine weevil feeding damage
The total amount of pine weevil feeding damage did not differ statistically significantly between soil classes (Table 2; p = 0.761). Pine weevils caused 19 and 33% of the mortality on peatland and mineral soil sites, respectively, without any statistically significant differences between the soil classes (p = 0.208).
The prediction models differed for the mineral soil and peatland. On peatland, the clear-cut age, site type, ground vegetation cover, type of planting spot and the interaction of the type of planting spot and the ground vegetation cover explained the probability of pine weevil feeding damage (Table 4). On mineral soil, the clear-cut age, ground vegetation cover and the type of planting spot explained the pine weevil feeding damage probabilities. When seedlings were planted on undisturbed soil there was always ground vegetation around the seedlings both on peatland and mineral soil.
| Table 4. The model for the pine weevil feeding damage in Norway spruce seedlings three years after planting. In the table, estimates, standard errors (SE) and the significance of parameters of fixed effects and the variances of random effects (geographical area, regeneration site and sample plot within a regeneration site) in a generalized linear mixed model are given. Soil classes [peatland (Peat) and mineral soil (Mineral)] were used as dummy variables and the fixed and random effect variances were estimated separately for each of them. In the final model, the fixed effects were clear-cut age (continuous), the site type (reference category “Sub-dry”), the ground vegetation cover around a seedling (reference category “No vegetation”) and the type of planting spot (reference category “Cultivated mineral soil”). CD is the coefficient of determination for different random effects. | ||||
| Effect type | Estimate (SE) | t Value | p-value | |
| Fixed effects | ||||
| Peat | 3.02 (1.93) | 1.56 | 0.144 | |
| Peat × Clear-cut age | –1.66 (0.40) | –4.12 | <0.001 | |
| Peat × Site type | Mesic | –2.29 (1.26) | –1.81 | 0.070 |
| Sub-mesic | 0.33 (0.40) | 0.81 | 0.418 | |
| Peat × Ground vegetation cover (V) | Dense | 2.91 (0.76) | 3.80 | <0.001 |
| Sparse | 1.85 (0.67) | 2.75 | 0.006 | |
| Peat × Type of planting spot (S) | Undisturbed soil (U) | 1.60 (0.62) | 2.58 | 0.010 |
| Cultivated organic (C) | 2.20 (0.68) | 3.24 | 0.001 | |
| Peat × V × S | U and dense | –0.59 (2.20) | –0.27 | 0.789 |
| C and dense | –2.18 (0.183) | –2.61 | 0.009 | |
| U and sparse | 0 | |||
| C and sparse | –0.92 (0.71) | –1.30 | 0.195 | |
| Mineral | 4.62 (1.35) | 3.41 | 0.006 | |
| Mineral × Clear-cut age | –1.81 (0.30) | –6.04 | <0.001 | |
| Mineral × Ground vegetation cover (V) | Dense | 2.65 (0.50) | 5.29 | <0.001 |
| Sparse | 1.31 (0.25) | 5.25 | <0.001 | |
| Mineral × Type of planting spot | Undisturbed soil | 1.11 (0.50) | 2.22 | 0.027 |
| Cultivated organic | 2.34 (0.33) | 7.01 | <0.001 | |
| Variances of random effects | Estimate (SE) | CD | ||
| Regeneration site (Peat) | 1.04 (0.50) | 0.54 | ||
| Sample plot (Peat) | 0.74 (0.23) | –0.15 | ||
| Regeneration site (Mine) | 0.46 (0.30) | 0.79 | ||
| Sample plot (Mine) | 0.90 (0.34) | –0.15 | ||
On mineral soil, when a site was planted one year after a clear-cut (clear-cut age ≤ 4 years), the feeding probability was over 0.1 independent of other variables (Fig 4a). When the seedlings were planted in cultivated mineral soil, the probability of feeding damage was lower than for other planting spots. Dense vegetation cover around a seedling increased the feeding damage probability. The probabilities were the highest, when the seedlings were planted in undisturbed soil or in cultivated planting spots covered by humus (organic material) and the seedlings were surrounded by dense vegetation. In those environments the probability of feeding damage was over 0.1, at six years after clear-cutting. On mineral soil, the feeding damage probabilities for seedlings planted in undisturbed soil surrounded by dense vegetation were 0.76 and 0.34 four and five years after clear-cutting.
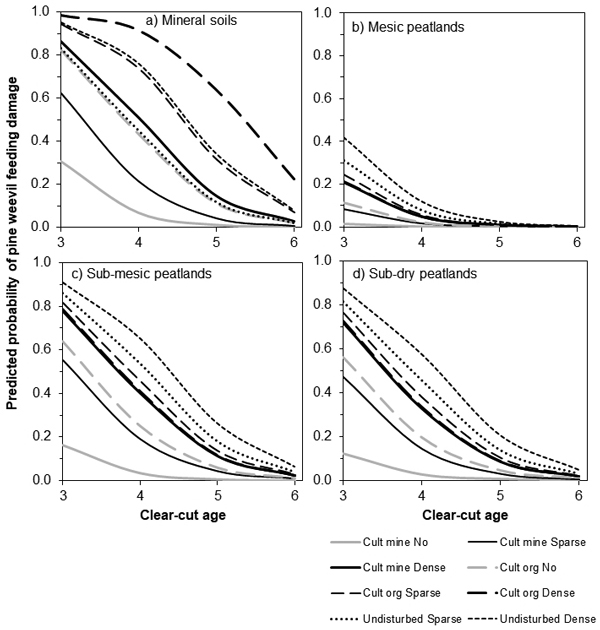
Fig. 4. The relationship of the clear-cut age and predicted probability of pine weevil feeding damage on Norway spruce seedlings in different classes in the ground vegetation cover and types of soil planting spots on (a) mineral soil and (b–d) peatland. On peatland, the predicted probabilities are presented also for (b) mesic, (c) sub-mesic and (d) sub-dry site types. In the data, there were no cases for undisturbed soil with no ground vegetation, thus those lines are not presented. Cult = cultivated (mound or patch) planting spot, Undisturbed = seedling planted in undisturbed soil, mine = planting spot covered by mineral soil, org = planting spot covered by peat in peatlands and humus in mineral soils.
On peatland, the site type clearly affected the feeding damage probability: the probability was lower on mesic sites than on other site types (Figs. 4b–d) independent of other variables. On all site types, the feeding damage probability was highest when a seedling was planted on undisturbed soil and it was surrounded by sparse or dense ground vegetation. When seedlings were planted in fresh clear-cuts (clear-cut age 3 years), the feeding damage probability was higher than for older clear-cuts, especially on sub-mesic and sub-dry sites (Figs. 4 c–d). The feeding damage probabilities for seedlings planted on undisturbed soil and surrounded by dense vegetation were 0.20–0.57 and 0.26–0.65 for four- and five-years old clear-cuts on sub-mesic and sub-dry sites, respectively. The corresponding values were 0.12 and 0.03 for mesic sites. The feeding damage probability was higher on cultivated peat (organic material) than on cultivated mineral soil when there was no or only sparse ground vegetation around a seedling. However, when a seedling was surrounded by dense vegetation, there were no differences between the cultivated planting spots. When a seedling was planted in cultivated mineral soil and there was no ground vegetation around a seedling, the feeding damage probability was less than 0.2 in all site types independent of the clear-cut age.
The model explained about 79% of the variation between regeneration sites on mineral soil sites and 54% on peatlands (Table 4). On the sample plot level, CDs were negative.
3.3.3 Ground vegetation cover
Three years after planting, 66% of seedlings were on spots where there was ground vegetation around them, but in most cases the vegetation cover was still classified as sparse (Table 2). The predicted probability for the presence of vegetation were 0.66 and 0.80 for mineral soil and peatland sites without any statistically significant differences between the soil classes (p = 0.088). However, the probability of dense vegetation cover was higher (p = 0.006) for peatland (0.06) than mineral soil (0.01).
On peatlands, the type of planting spot, the site type and the MSP method and interaction of the site type and MSP method with the planting spot predicted the presence of ground vegetation three years after planting (Table 5; Fig. 5). On mesic sites and on undisturbed soil the probability of vegetation was always 1.0. On sub-mesic sites the probability was higher than on sub-dry sites both on cultivated peat and in cultivated mineral soil. Differences in the probability of vegetation due to different MSP methods were small on cultivated planting spots, although the trend was that ground vegetation was more probable for spot mounding than in other MSP methods on all cultivated spots, and more probable on cultivated peat than mineral soil for spot mounds with no differences between cultivated spots with soil inversion or ditch mounding.
| Table 5. The model for the presence of ground vegetation around a Norway spruce seedling three years after planting. Estimates, standard errors (SE) and the significance of parameters of fixed effects and the variances of random effects (regeneration site and sample plot within a regeneration site) in a generalized linear mixed model are given. Soil classes [peatland (Peat) and mineral soil (Mineral)] were used as dummy variables and the fixed and random effect variances were estimated separately for each of them. In the final model, fixed effects were the clear-cut age (continuous), the site type (reference category “Sub-dry”), the mechanical site preparation (MSP) method (reference category “Spot mounding”) and the type of planting spot (reference category “Cultivated mineral soil”). CD is the coefficient of determination for different random effects. | ||||
| Effect type | Estimate (SE) | t Value | p-value | |
| Fixed effects | ||||
| Peat | 1.21 (0.66) | 1.83 | 0.090 | |
| Peat × Site type (T) | Mesic (OMT) | 14.77 (476) | 0.03 | 0.975 |
| Sub-mesic (MT) | –0.16 (0.54) | –0.29 | 0.769 | |
| Peat × MSP | Soil inversion (I) | –0.28 (0.76) | –0.36 | 0.717 |
| Ditching and mounding (D) | –0.26 (0.72) | –0.36 | 0.716 | |
| Peat × Type of planting spot (S) | Undisturbed soil (U) | 31.59 (0) | Infty | <0.001 |
| Cultivated peat (C) | 0.47 (0.65) | 0.71 | 0.476 | |
| Peat × T × S | OMT and C | –9.93 (476) | –0.02 | 0.983 |
| MT and U | 23.14 (0) | Infty | <0.001 | |
| MT and C | 1.49 (0.56) | 2.64 | 0.008 | |
| Peat × MSP × S | I and U | 5.46 (0) | Infty | <0.001 |
| I × C | –1.07 (0.75) | –1.43 | 0.152 | |
| D × U | 12.04 (0) | Infty | <0.001 | |
| D × C | –1.21 (0.79) | –1.53 | 0.125 | |
| Mineral | –1.11 (0.91) | –1.22 | 0.249 | |
| Mineral × Clear-cut age (A) | 0.33 (0.19) | 1.72 | 0.086 | |
| Mineral × Type of planting spot (S) | Undisturbed soil (U) | 25.74 (0) | Infty | <0.001 |
| Cultivated humus (C) | 0.92 (0.27) | 3.39 | <0.001 | |
| Variances of random effects | Estimate (SE) | CD | ||
| Regeneration site (Peat) | 1.10 (0.52) | 0.41 | ||
| Sample plot (Peat) | 1.24 (0.31) | 0.16 | ||
| Regeneration site (Mine) | 0.16 (0.12) | 0.60 | ||
| Sample plot (Mine) | 0.87 (0.22) | –0.07 | ||
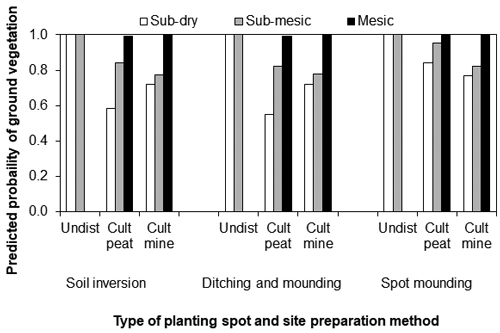
Fig. 5. The predicted probability that there would be ground vegetation cover around a Norway spruce seedling three years after the planting for different types of sites, planting spots and mechanical site preparation methods on peatland sites. In the data, there were no cases where seedlings had been planted on undisturbed soil on mesic sites, thus those bars are not presented. Cult = cultivated (mound or patch) planting spot, Undist = a seedling planted in undisturbed soil, mine = planting spot covered by mineral soil, peat = planting spot covered by peat.
On mineral soil, the age of a clear-cut and the type of the planting spot explained the presence of ground vegetation around a seedling (Table 5). We included the clear-cut age in the model although it was nearly significant. By including it, the CD of the model increased at the regeneration site level. When a seedling was planted in undisturbed soil, the probability of surrounding vegetation was always 1.0 independent of the clear-cut age (Fig. 6). On cultivated humus, the probability of surrounding vegetation was always > 0.68, increasing with the age of the clear-cuts. On cultivated mineral soil, the presence of ground vegetation was more probable the older the clear-cut was.
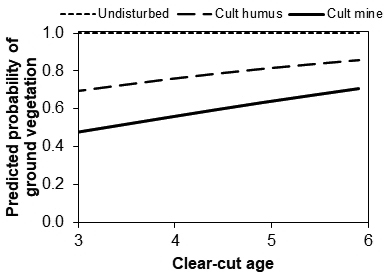
Fig. 6. The effect of clear-cut age on the predicted probability that there would be ground vegetation cover around a Norway spruce seedling three years after planting in different types of planting spots in mineral soils.
4 Discussion
4.1 Planting success
Our results provide novel information about the reasons for differences in the planting success on mineral soil and peatland sites in boreal forest conditions. The results indicate that the plating success is poorer on peatlands than on mineral soil and is in accordance with previous studies. In the study by Kankaanhuhta et al. (2009) a reduction in the number of seedlings for the “Peat” soil class was 10% compared to mineral soils with a medium coarse texture. In their study, the planting success was poorer than in our study, while the average number of planted seedlings for all soil texture types was similar (1388 seedlings ha–1) to the average number of planted seedlings of peatland sites in our study. Thus, compared to the average planting success in privately owned forests in Finland at the beginning of the 2000s in the study by Kankaanhuhta et al. (2009), the planting success in our study was clearly better on mineral soil. Compared to the machine planting survey by Luoranen et al. (2011), the average number of surviving seedlings per ha–1 on mineral soil sites was approximately the same level, with 1600 seedlings ha–1.
The poorer planting success on peatland sites was mainly caused by differences in the ground vegetation cover surrounding the seedlings since dense vegetation cover was more probable on peatland than on mineral soil. The dense ground vegetation cover increased the risk of pine weevil feeding damage, as was also shown by Petersson et al. (2006). Additionally, dense vegetation cover may also increase the risk of vole damage (Teivainen et al. 1986; Huitu et al. 2013). In our study, most of the pine weevil feeding damage was minor with no effect on the seedling survival. This was an inventory study, in which planting areas were measured three years after cultivation. Seedlings were also not marked when planting. Thus, our survival and damage estimates are probably underestimated, since all planted but later dead seedlings were unlikely to be found at the time of measurement. The survival of the seedlings was highest on mineral soil covered mounds and more cultivated mineral soils were found on mineral soils than on peatland sites. This is in accordance with the results by Åkerström and Hånell (1997), who observed the increased survival of seedlings on mineral soil-covered mounds on peatland. On the other hand, the soil texture type in the mounds on peatland affects the development of the ground vegetation. The development is the fastest on mounds with a mixture of mineral soil (Saarinen et al. 2009). The ground vegetation cover, the type of planting spot and the probability of pine weevil feeding damage are linked and may explain the differences in the planting success on peatland and mineral soil.
4.2 Ground vegetation
Our results showed that the cover of vegetation around a seedling on peatland depended on the site type and was more common on mesic-peatland sites than other site types. This is in accordance with previous studies. In the study by Saarinen et al. (2009), the vegetation development was slower on less fertile peatland than on fertile types of peatland. In our study, mineral soil sites were all sub-mesic and sub-dry, which are less fertile than peatland sites.
The lower probability of dense ground vegetation growing around seedlings on mineral soil indicates that the development of vegetation was slower on mineral soil than on peatland, especially on cultivated planting spots. Previously, the increasing cover of ground vegetation around seedlings on older regeneration sites has been also observed by Örlander et al. (1996) and Hanssen et al. (2003) on mineral soil in South Sweden and Norway. Örlander et al. (1996) also found that the vegetation development was faster on unprepared soil than on mounds. Furthermore, Saksa (2013) observed that the number of birch seedlings increased when the soil texture type was peat in comparison to medium coarse soils or if the main species in the ground layer consisted of Sphagnum or other mosses. In our study, there was always ground vegetation in the vicinity of a seedling if a seedling was planted in undisturbed soil both on peatland and mineral soil. This is in accordance with the results by Hallsby (1995) that the development of herbaceous vegetation was faster on undisturbed soil or with humus-cover than on bare mineral soil.
When the seedlings were planted in spot mounds, there was slightly more ground vegetation around the seedling than for other types of mounds on peatland. In spot mounding, more soil is disturbed as there is both a mound and a patch beside it, while in other methods, the soil is inverted on the same spot, or the soil is dropped onto undisturbed soil. Disturbed soil is a better surface for the emergence of seed-born broadleaves. Laine et al. (2020) observed that there was a slightly larger area of cultivated mineral soil and more naturally regenerated birches per hectare in spot mounded areas than in areas where the inverted method had been used on mineral soil sites in Central Finland.
4.3 Pine weevil feeding damage
The probability of pine weevil feeding damage modeled in this study predicts the occurrence of any level of pine weevil damage, including small feeding scars without any effect on the seedling health. In our survey, some level of pine weevil damage was found for all the inventoried seedlings, including the seedlings which died in previous years. Thus, the prediction model describes the probability of the overall damage caused by pine weevil feeding from planting until the inventory, not solely the damage in the year of inventory. There were also planting spots without seedlings (2–3% of mounds) and a part of these seedlings had probably died due to pine weevil feeding, so the prediction model may underestimate the total probability of feeding damage. The levels of pine weevil feeding damage did not differ between peatland and mineral soil sites. However, different factors predicted the feeding damage on different soil classes.
4.3.1 Clear-cut age
The clear-cut age at the time of planting affected the probability of pine weevil feeding damage even after three years from planting. On mineral soil, the effect of the clear-cut age on pine weevil feeding damage has been observed in several previous studies (Luoranen et al. 2017; Nordlander et al. 2017). The feeding damage probability of seedlings planted on undisturbed soil indicates the size of the pine weevil population on the regeneration site, i.e., the feeding pressure (Luoranen et al. 2017). On mineral soil sites, there is a risk for pine weevil damage at least two years after clear-cutting. On peatland, it seemed that the probability of feeding damage decreased earlier than on mineral soil sites, especially on mesic sites. However, this probably underestimated the cumulative feeding damage from the time of planting until the inventory. On peatland, the probability of survival was lower when there was dense vegetation cover surrounding the seedlings. On mesic peatland sites, there was always vegetation around the seedlings. Thus, it is probable that we did not find all the seedlings that had died in the earlier years, and for which we could not determine the cause of death. Previously, Långström (1982) observed high pine weevil population levels at least four years after clear-cuts in Central-Finland.
On cultivated mineral soil, the effect of the clear-cut age on the probability of pine weevil feeding damage was approximately the same for similar site types on peatland and mineral soil. Despite the higher feeding pressure on mineral soil sites for younger clear-cuts, the protection effect of soil cultivation, especially with mineral soil cover, was the same for both soil classes independent of the clear-cut age. The feeding damage probabilities for cultivated mineral soil without ground vegetation on mesic sites on peatland in our study were approximately the same as observed by Piri et al. (2020) on clear-cuts with a corresponding age (three- or four-year-old) for stump removal sites in South and Central-Finland. For the other cases, the feeding damage probabilities in our study were much higher. In the study by Piri et al. (2020), only the current year’s feeding damage was included in the prediction model while we included the cumulative damage between planting and inventory.
The survey conducted one growing season after planting discovered that pine weevil feeding damage decreased with the age of the clear-cut (Luoranen and Viiri 2012). In this study, taking place three years after planting, a similar trend was observed. Although pine weevil feeding does not kill the seedlings, it may reduce their growth (Luoranen et al. 2017) and poorly growing seedlings are more susceptible to ground vegetation competition.
4.2.2 Site type
Based on the model, the soil fertility affected the feeding damage probability substantially and was much lower on mesic sites than less fertile and drier peatland site types. On mineral soil, the site type was not a statistically significant predictor in the model. This was probably caused by the small number (peatland) or lack (mineral soil) of the most fertile sites in the data and the result is only indicative. Even so, our results considering peatland are in accordance with previous results by Långström (1982). He found more pine weevils on dry, pine-dominated sites compared with mesic, spruce-dominated sites. Despite the differences in the feeding damage probability between the site types, the site type did not affect the survival of the seedlings in our study.
4.2.3 Ground vegetation and pine weevils
The probability of pine weevil feeding damage increased with increasing ground vegetation cover around a seedling both on peatland and mineral soil sites. Surrounding ground vegetation, even when it does not grow on the mound itself, creates more favorable conditions for pine weevils (Petersson et al. 2006), and increases pine weevil damage (Örlander and Nordlander 2003; Petersson and Örlander 2003; Petersson et al. 2006). This was also the case in the present study, as the increasing cover of vegetation was found to increase the feeding damage probability regardless of the type of planting spot. The difference was greater between vegetation classes on cultivated mineral soil than on cultivated peat on peatland. This disagrees with the results by Petersson et al. (2006), who observed that the surrounding vegetation did not affect the pine weevil feeding on humus covered planting spots in the first season. The differences between the studies can probably be explained by the time between planting and the inventory, as well as the different definitions of the surrounding vegetation. Our study was done three years after planting and the ground vegetation cover was assessed in the immediate surrounding of the seedling, whereas in the study by Petersson et al. (2006) the vegetation grew outside the planting spot and gave shelter above seedlings.
4.2.4 The type of planting spot
The MSP method did not affect the survival of the seedlings or pine weevil feeding damage for either of the soil classes. The quality of MSP and the type of the planting spot were more important (Wallertz et al. 2018). Mineral soil cover around a seedling is known to reduce the probability of pine weevil feeding damage (Björklund et al. 2003; Luoranen et al. 2017) as was found in our study. There was less feeding damage on mineral soil-covered planting spots than on other types of planting spots, both on peatland and mineral soil sites. On peatland, mineral soil cover in the mounds was possible since all the peatland sites were drained and had a very thin peat layer. The third-year result was in accordance with a survey done after the first growing season (Luoranen and Viiri 2012): the probability of damage was clearly higher on peat and humus-covered spots compared to spots with mineral soil. Our result is also in accordance with several previous studies (e.g., Nordlander et al. 2011; Pearson et al. 2011; Sikström et al. 2020). In the study by Hytönen et al. (2020), pine weevil damage was found in planted Scots pine (Pinus sylvestris L.) seedlings on unprepared soil but not on prepared soil on drained peatland in Northern Finland, where the pine weevil feeding pressure is lower than in South and Central Finland. Our results on peatland sites are in accordance with the study by Pearson et al. (2011) who found more pine weevil damage on unprepared or scalped planting spots than for mounds on deep-peated regeneration sites in Central Finland.
4.3 Statistical inference
The data for this study came from an inventory of practical regeneration sites planted in one year, not from a designed experiment. The amount of data was quite small, with only 30 sites located in Central Finland and the models are valid for the corresponding boreal conditions. Different companies operated in the geographical areas (regions) and this affected the study. For instance, each company had its own guidelines on MSP methods and how many years there had to be between clear-cutting and planting. In addition, there were no mesic mineral soil sites and only a few mesic peatland sites. Thus, one needs to be careful when making causal inferences from the statistical relations. In some cases, there were negative CD-values at the sample plot level. This is probably caused by estimation errors.
5 Conclusions
On peatland, there were a high proportion of seedlings planted on organic-covered planting spots and surrounded by ground vegetation three years after planting. Both these factors increased the probability of pine weevil feeding damage and reduced the probability of seedling survival. On mineral soil sites, the seedlings were mainly planted in cultivated mineral soil and the probability of harmful vegetation around a seedling was more probable on older (five to six years after planting) regeneration sites. Due to these differences, the planting success of Norway spruce seedlings was poorer on peatland sites compared to the mineral soil sites. To ensure the proper planting success on peatland regeneration sites, the quality of the soil preparation and control of the competing vegetation during the first years after planting should be emphasized, especially on the fertile site types typical to Norway spruce plantings. Additionally, on peatland regeneration sites with a thin peat layer, the soil preparation method selected should aim to maximize the amount of mineral soil covered mounds, as this promotes seedling survival. These results are not valid for sites with a thick peat layer and planting success or alternative methods for artificial regeneration on those sites should be further studied.
Authors’ contributions
Both authors contributed to the conception of the research questions and design of the work. JL analyzed the data. Both authors participated in the interpretation of data and results, and writing.
Acknowledgements
This study was conducted in cooperation with Metsähallitus, Metsäliitto Osuuskunta and UPM Forest. We thank them for their cooperation. We are grateful to Pekka Rossi for the field data collection and Liisa Kylmälä for the data processing for analysis. We thank Dr. Päivi Väänänen for the valuable comments on the manuscript. The language was checked by Acolad (previously AAC Global). This work was supported by previous Finnish Forest Research Institute and Natural Resources Institute Finland (Projects 3554, 41007-0012500, 41007-00100100).
Declaration of openness of research materials, data, and code
Collected inventory data are available upon request from corresponding author (jaana.luoranen@luke.fi).
References
Åkerström L, Hånell B (1997) Mound characteristics affect growth and survival of Norway spruce seedlings. In: Trettin CC, Jurgensen MF, Grigal DF, Gale MR, Jeglum JK (eds) Northern forested wetlands and management. Boca Raton, Flo, CRC Lewis, pp. 429–435. https://doi.org/10.1201/9780203745380-30.
Björklund N, Nordlander G, Bylund H (2003) Host‐plant acceptance on mineral soil and humus by the pine weevil Hylobius abietis (L.). Agric For Entomol 5: 61–65. https://doi.org/10.1046/j.1461-9563.2003.00163.x.
Cajander AK (1949). Forest types and their significance. Acta For Fenn 56, article id 7396. https://doi.org/10.14214/aff.7396.
Hallsby G (1995) Field performance of outplanted Norway spruce: effects of organic matter amendments and site preparation. Can J For Res 25: 1356–1367. https://doi.org/10.1139/x95-148.
Hånell B (1993) Regeneration of Picea abies forests on highly productive peatlands – clear-cutting or selective cutting? Scand J For Res 8: 518–527. https://doi.org/10.1080/02827589309382798.
Hanssen KH, Granhus A, Brække FH, Haveraaen O (2003) Performance of sown and naturally regenerated Picea abies seedlings under different scarification and harvesting regimens. Scand J For Res 18: 351– 361. https://doi.org/10.1080/02827580310005973.
Holgén P, Hånell B (2000) Performance of planted and naturally regenerated seedlings in Picea abies-dominated shelterwood stands and clearcuts in Sweden. For Ecol Manage 127: 129–138. https://doi.org/10.1016/S0378-1127(99)00125-5.
Huitu O, Rousi M, Henttonen H (2013) Integration of vole management in boreal silvicultural practices. Pest Manage Sci 69: 355–361. https://doi.org/10.1002/ps.3264.
Hytönen J, Hökkä H, Saarinen M (2020) The effect of planting, seeding and soil preparation on the regeneration success of Scots pine (Pinus sylvestris L.) on drained peatlands – 10-year results. Forestry Studies | Metsanduslikud Uurimused 72: 91–106. https://doi.org/10.2478/fsmu-2020-0008.
Kankaanhuhta V, Saksa T, Smolander H (2009) Variation in the results of Norway spruce planting and Scots pine direct seeding in privately-owned forests in southern Finland. Silva Fenn 43: 51–70. https://doi.org/10.14214/sf.217.
Korhonen KT, Ihalainen A, Ahola A, Heikkinen J, Henttonen HM, Hotanen J-P, Nevalainen S, Pitkänen J, Strandström M, Viiri H (2017) Suomen metsät 2009–2013 ja niiden kehitys 1921–2013. [Forests in Finland 2009–2013 and the development of them 1921–2013]. Luonnonvara- ja biotalouden tutkimus 59/2017. Luonnonvarakeskus, Helsinki. http://urn.fi/URN:ISBN:978-952-326-467-0.
Laine T, Kankaanhuhta V, Rantala J, Saksa T (2020) Effects of spot mounding and inverting on growth of conifers, exposed mineral soil and natural birch regeneration. Silva Fenn 54, article id 10369. https://doi.org/10.14214/sf.10369.
Långström B (1982) Abundance and seasonal activity of adult Hylobius-weevils in reforestation areas during first years following final felling. Comm Inst Fenn 106. http://urn.fi/URN:ISBN:951-40-0575-9.
Luoranen J, Viiri H (2012) Soil preparation reduces pine weevil (Hylobius abietis L.) damage on both peatland and mineral soil sites one year after planting. Silva Fenn 46: 151–161. https://doi.org/10.14214/sf.71.
Luoranen J, Rikala R, Smolander H (2011) Machine planting of Norway spruce by Bracke and Ecoplanter: an evaluation of soil preparation, planting method and seedling performance. Silva Fenn 45: 341–357. https://doi.org/10.14214/sf.107.
Luoranen J, Viiri H, Sianoja M, Poteri M, Lappi J (2017) Predicting pine weevil risk: effects of site, planting spot and seedling level factors on weevil feeding and mortality of Norway spruce seedlings. For Ecol Manage 389: 260–271. https://doi.org/10.1016/j.foreco.2017.01.006.
Luoranen J, Saksa T, Lappi J (2018) Seedling, planting site and weather factors affecting the success of autumn plantings in Norway spruce and Scots pine seedlings. For Ecol Manage 419–242: 79–90. https://doi.org/10.1016/j.foreco.2018.03.040.
Moilanen M, Ferm A, Issakainen I (1995) Kuusen- ja koivuntaimien alkukehitys korven uudistusaloilla. [Development of Norway spruce and birch seedlings in regeneration on spruce mires]. Metsätieteen aikakauskirja 2/1995: 115–130. https://doi.org/10.14214/ma.5950.
Nieminen M, Hökkä, H, Laiho R, Juutinen A, Ahtikoski A, Pearson M, Kojola S, Sarkkola S, Launiainen S, Valkonen S, Penttilä T, Lohila A, Saarinen M, Haahti K, Mäkipää R, Miettinen M, Ollikainen M (2018) Could continuous cover forestry be an economically and environmentally feasible management option on drained boreal peatlands? For Ecol Manage 424: 78–84. https://doi.org/10.1016/j.foreco.2018.04.046.
Nilsson U, Örlander G (1995) Effects of regeneration methods on drought damage to newly planted Norway spruce seedlings. Can J For Res 25: 790–802. https://doi.org/10.1139/x95-086.
Nordlander G, Hellqvist C, Johansson K, Nordenhem H (2011) Regeneration of European boreal forests: effectiveness of measures against seedlings mortality caused by the pine weevil Hylobius abietis. For Ecol Manage 262: 2354–2363. https://doi.org/10.1016/j.foreco.2011.08.033.
Nordlander G, Mason EG, Hjelm K, Nordenhem H, Hellqvist C (2017) Influence of climate and forest management on damage risk by pine weevil Hylobius abietis in northern Sweden. Silva Fenn 51, article id 7751. https://doi.org/10.14214/sf.7751.
Örlander G, Nordlander G (2003) Effects of field vegetation control on pine weevil (Hylobius abietis) damage to newly planted Norway spruce seedlings. Ann For Sci 60: 667–671. https://doi.org/10.1051/forest:2003059.
Örlander G, Nilsson U, Hällgren J-E (1996) Competition for water and nutrients between ground vegetation and planted Picea abies. New Zeal J For 26: 99–117. https://www.scionresearch.com/__data/assets/pdf_file/0004/59557/NZJFS261-21996ORLANDER99-117.pdf.
Pearson M, Saarinen M, Minkkinen K, Silvan N, Laine J (2011) Mounding and scalping prior to reforestation of hydrologically sensitive deep-peated sites: factors behind Scots pine regeneration success. Silva Fenn 45: 647–667. https://doi.org/10.14214/sf.98.
Petersson M, Örlander G (2003) Effectiveness of combination of shelterwood, scarification, and feeding barriers to reduce pine weevil damage. Can J For Res 33: 64–73. https://doi.org/10.1139/x02-156.
Petersson M, Nordlander G, Örlander G (2006) Why vegetation increases pine weevil damage: bridge or shelter? For Ecol Manage 225: 368–377. https://doi.org/10.1016/j.foreco.2006.01.012.
Piri T, Viiri H, Hyvönen J (2020) Does stump removal reduce pine weevil and other damage in Norway spruce regeneration? – Results of a 12-year monitoring period. For Ecol Manage 465, article id 118098. https://doi.org/10.1016/j.foreco.2020.118098.
Saarinen M (2013) Artificial and natural seeding of Scots pine in old drainage areas – unique features of forest regeneration on peatlands. Diss For 164. https://doi.org/10.14214/df.164.
Saarinen M, Hotanen J-P, Alenius V (2009) Vegetation succession in prepared microsites in drained peatland forest regeneration areas. Suo 60: 95–109. http://www.suo.fi/pdf/article9869.pdf.
Saksa T (2013) Regeneration after stump harvesting in southern Finland. For Ecol Manage 219: 79–82. https://doi.org/10.1016/j.foreco.2012.08.014.
Sikström U, Hjelm K, Holt Hanssen K, Saksa T, Wallertz K (2020) Influence of mechanical site preparation on regeneration success of planted conifers in clearcuts in Fennoscandia – a review. Silva Fenn 54, article id 10172. https://doi.org/10.14214/sf.10172.
Silver T, Piri T (2017) Observations of Heterobasidion root rot in Scots pine stands on peatlands in southern Finland Suo 68: 1–12. http://www.suo.fi/pdf/article10110.pdf.
Teivainen T, Jukola-Sulonen E-L, Mäenpää E (1986) The effect of ground vegetation suppression using herbiside on the field vole, Migrotus agretis (L.) population. Folia For 651. http://urn.fi/URN:ISBN:951-40-0734-4.
Tonteri T, Hotanen J-P, Kuusipalo J (1990) The Finnish forest site type approach: ordination and classification studies of mesic forest sites in southern Finland. Vegetatio 87: 85–98. https://doi.org/10.1007/BF00045658.
Vasander H, Tuittila E-S, Lode E, Lundin L, Ilomets M, Sallantaus T, Heikkilä R, Pitkänen M-L, Laine J (2003) Status and restoration of peatlands in northern Europe. Wet Ecol Manage 11: 51–63. https://doi.org/10.1023/A:1022061622602.
Viiri H, Tuomainen A, Tervo L (2007) Persistence of deltamethrin against Hylobius abietis on Norway spruce seedlings. Scand J For Res 22: 128–135. https://doi.org/10.1080/02827580701224113.
Wallertz K, Björklund N, Hjelm K, Petersson M, Sundblad L-G (2018) Comparison of different site preparation techniques: quality of planting spots, seedlings growth and pine weevil feeding damage. New For 49: 705–722. https://doi.org/10.1007/s11056-018-9634-8.
Total of 38 references.
