Āris Jansons,
Roberts Matisons  ,
Līga Puriņa,
Una Neimane,
Jānis Jansons
,
Līga Puriņa,
Una Neimane,
Jānis Jansons
Relationships between climatic variables and tree-ring width of European beech and European larch growing outside of their natural distribution area
Jansons Ā., Matisons R., Puriņa L., Neimane U., Jansons J. (2015). Relationships between climatic variables and tree-ring width of European beech and European larch growing outside of their natural distribution area. Silva Fennica vol. 49 no. 1 article id 1255. https://doi.org/10.14214/sf.1255
Highlights
- In western Latvia, variation of tree-ring width of European beech and European larch within stands was similar
- Dry summers and cold winters caused common event years in tree-rings
- Moisture availability at the end of summer was apparently the main limiting factor for tree-ring width
- Winter and spring temperature did not have significant and lasting effect on variation of tree-ring width.
Abstract
Relationships between climatic variables and tree-ring width (TRW) of dominant European larch (Larix decidua Mill.) and European beech (Fagus sylvatica L.) trees growing outside of their natural distribution area in western Latvia were studied. Chronologies of TRW, which covered the periods 1949–2012 and 1911–2012, were produced for beech and larch, respectively. Common signatures in TRW between both species were observed, but their amplitude differed. Correlation analysis showed that variation of TRW of both species was affected by drought related climatic variables. Tree-ring width of beech was affected by temperature in the previous July and August and the effect of spring and autumn temperature was observed. Since the 1980s, the effect of July precipitation has become significant. Summer precipitation was significant for larch in the mid-part of the previous century; however, temperature in the previous September has become a limiting factor since 1970s. The limiting effect of winter and spring temperature apparently lost its significance around the 1950s.
Keywords
Fagus sylvatica;
introduced species;
wood increment;
climatic limitation;
Latvia;
Larix decidua
- Jansons, LSFRI „SILAVA”, Rigas Str. 111, Salaspils, Latvia, LV2169 E-mail aris.jansons@silava.lv
-
Matisons,
LSFRI „SILAVA”, Rigas Str. 111, Salaspils, Latvia, LV2169
E-mail
robism@inbox.lv
- Puriņa, LSFRI „SILAVA”, Rigas Str. 111, Salaspils, Latvia, LV2169 E-mail liga.purina@silava.lv
- Neimane, LSFRI „SILAVA”, Rigas Str. 111, Salaspils, Latvia, LV2169 E-mail una.neimane@silava.lv
- Jansons, Forest Competence Centre, Dzerbenes str. 27, Riga, Latvia, LV1006 E-mail janis.jansons@mnkc.lv
Received 24 September 2014 Accepted 9 January 2015 Published 23 January 2015
Views 97490
Available at https://doi.org/10.14214/sf.1255 | Download PDF

1 Introduction
Climatic changes cause shifts in distribution of tree species (Walther et al. 2002; Kullman 2008) and affect forest productivity (Lindner et al. 2010). Although warming of climate might increase productivity of boreal forests (Lindner et al. 2010), changes in forest composition are also predicted (Hickler et al. 2012). In this regard, adjustments in forest management practices are crucial to ensure sustainable forestry. One such adjustment may be the introduction of novel species, however, their ecological demands, i.e. climatic limitation of growth, should be comprehensively evaluated. Detailed information on climate-growth relationships can be obtained via dendrochronological analysis, especially for trees growing close to their natural distribution limit or outside of it (Fritts 2001; Vetaas 2002).
In Latvia, European beech (Fagus sylvatica L.) and European larch (Larix decidua Mill.) occur northwards from their natural distribution area (Bolte et al. 2007; EUFORGEN 2009) and cover 43 and 1139 ha of forestlands, respectively; both of these species are productive (Dreimanis 1995). Although warming of climate appears beneficial for growth of these species in Latvia (Hickler et al. 2012), the effect of climatic variables might still be substantial (Fritts 2001). The aim of this study was to assess the relationships between climatic variables and tree-ring width (TRW) of larch and beech in western Latvia. We hypothesized that winter-spring temperature has been limiting TRW, but that these effects have weakened during the 20th century.
2 Material and methods
2.1 Study area, sampling and measurements
European larch and European beech growing in western Latvia in even-aged plantations near Auce (56°31´N, 22°56´E, 50 m a.s.l.) and Kaleti (56°21´N, 21°29´E, 80 m a.s.l.), respectively, were studied. North-eastern Germany is considered as the area of origin for both species. Mean dimensions of the studied trees are shown in Table 1. Plantations were situated on flat terrain on fertile, well-drained clayey soils. Climate in the study areas is mild: mean annual temperature is ca.+6.1 °C, July is the warmest month with a mean temperature ca. +16 °C, and January is the coldest month with a mean temperature ca. –3.6 °C. The period when the mean daily temperature exceeds + 5 °C is ca. 185 days. Annual precipitation sum is ca. 560 and ca. 750 mm in the Auce and Kaleti sites, respectively. Most of the precipitation falls during summer. Climatic changes are reflected as an increase of temperature in the dormant period and spring, during the 20th century, mean November–April temperature has increased by 0.83 and 1.10 °C in Kaleti and Auce, respectively, and an intensification of heat and drought events (Avotniece et al. 2010).
| Table 1. Mean dimensions of sampled trees and statistics of TRW (tree-ring width) datasets: number of measured and crossdated time-series; range, agreement (interseries correlation, first order autocorrelation, GLK (Gleichläufigkeit) and EPS (expressed population signal)) and mean sensitivity of crossdated tree time-series of beech and larch. | ||
| Beech | Larch | |
| Studied period | 1949–2012 | 1911–2012 |
| Number of samples | 18 | 44 |
| Number of crossdated samples | 18 | 38 |
| Number of crossdated trees | 9 | 20 |
| Mean tree height, m | 28.3 | 32.2 |
| Standard deviation of tree height, m | 4.26 | 3.17 |
| Mean tree diameter at breast height, cm | 41.5 | 48.7 |
| Standard deviation of diameter at breast height, cm | 6.3 | 10.7 |
| Min TRW, mm | 0.30 | 0.23 |
| Max TRW, mm | 6.63 | 13.22 |
| Mean TRW, mm | 2.80 | 2.40 |
| Standard deviation of TRW, mm | 1.01 | 1.62 |
| Interseries correlation | 0.69 | 0.60 |
| First order autocorrelation | 0.61 | 0.76 |
| Mean sensitivity | 0.22 | 0.29 |
| GLK | 0.70 | 0.64 |
| EPS | 0.88 | 0.91 |
In the studied sites, visually healthy, dominant trees, 22 larches and 9 beeches, were cored at breast height. Two samples per tree from opposite sides of stem were collected using a Pressler corer. In the laboratory, air-dried cores were fixed and gradually grinded (sandpaper roughness 100, 150, 250 and 400 grains per inch). Tree-ring width was measured using the LINTAB 5 (RinnTECH, Heidelberg, Germany) measurement system with precision of 0.01 mm.
2.2 Data analysis
Time series of TRW were cross-dated and their quality was checked graphically and using the program COFECHA (Grissino-Mayer 2001). Mean time series of TRW for trees were produced and rechecked. Residual chronologies of TRW were produced using the program ARSTAN (Cook and Holmes 1986); double detrending with negative exponential curve, cubic-spline with 128-year wavelength and autoregressive modelling were used. Statistics of datasets were calculated using library “dplR” in R (Bunn 2008). Chronologies were compared using the Gleichläufigkeit (GLK) and Pearson correlation coefficients. Gridded climatic data: mean monthly temperature and precipitation sums were obtained from the Climatic Research Unit (Jones et al. 1999) for points closest to the sites. The effect of climatic variables on TRW was assessed by bootstrapped Pearson correlation and response function analysis using the program DendroClim2002 (Biondi and Waikul 2004) for the whole chronologies and by 40-year moving intervals. Time windows from June of the preceding year to October of the current year were used for climatic data.
3 Results
Most of the measured time series of TRW (Fig. 1 A,B) were maintained for production of the chronologies. Time series of TRW showed good agreement as EPS were above 0.85 and interseries correlation were above 0.60 (Table 1), thus confirming the validity of datasets. Considering that autocorrelation was high (Table 1), residual chronologies were produced (Fig. 1 C). Chronologies were synchronous, as GLK was 0.59, but the correlation coefficient between them was rather low (r = 0.27, p-value = 0.03). Common decreases of TRW were observed in 1979, 1992, 1995, 1998, 2006 and 2008, and common increases were observed in 1957, 1978 and 1994 (Fig. 2 C).
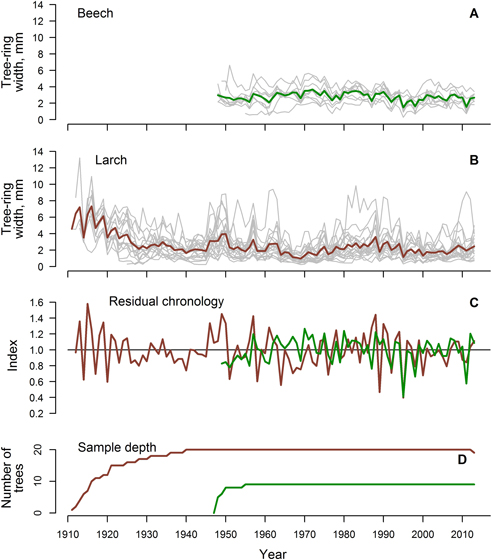
Fig. 1. Crossdated time series of TRW of beech (A) and larch (B), residual chronologies (C) and sample depth (D) (in C and D, green line represents beech and red line represents larch).
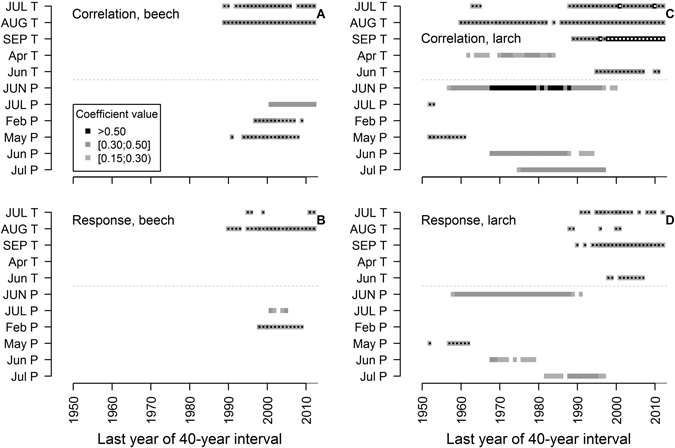
Fig. 2. Significant (p-value < 0.05) Pearson correlation (black bars) and response function (grey bars) coefficients between climatic variables: mean monthly temperature (T) and precipitation sums (P) and residual chronologies of beech (A) and larch (B) TRW, calculated using data for whole period (1911–2012 for larch and 1949–2012 for beech). Months in uppercase correspond to the year prior to tree-ring formation (t–1).
The established chronologies showed significant correlation (|r| < 0.41) and/or response function with seven of the tested 34 climatic variables, when the entire periods were analysed (Fig. 2). Tree-ring width of beech was mainly affected by temperature in the previous July and August as suggested by the significance of both coefficients. Temperature in the previous October and current May also showed a significant effect on TRW of beech. The analysis of moving intervals showed (Fig. 3 A,B) that temperature in the previous August has had a stable effect throughout the analysed period, while the effect of precipitation in the previous July has become significant in moving intervals after 1960–2000. Precipitation in February and May has been significant for TRW in the mid part of the analysed period (Fig. 3 A,B).
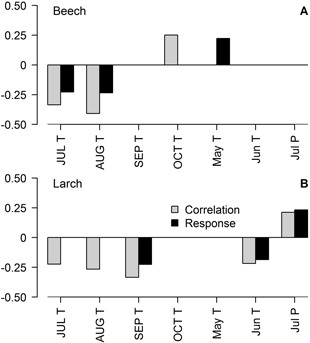
Fig. 3. Significant (p-value < 0.05) Pearson correlation and response function coefficients between climatic variables: mean monthly temperature (T) and precipitation sums (P) and residual chronologies of beech TRW (A and B, respectively) and larch (C and D, respectively) calculated for 40-year moving intervals. Months in uppercase correspond to the year prior to tree-ring formation (t–1). Dots represent negative coefficient values. Note that length of the analysed chronologies differs between species.
When the entire larch chronology was analysed, temperature in the previous September and current June and precipitation in July were the main limiting variables for TRW (Fig. 2 B), as suggested by correlation and response coefficients, although coefficient values were lower compared to beech (Fig. 2 A). Nevertheless, TRW of larch was also correlated with temperature in the previous July and August, similarly to beech (Fig. 2 A). Changes in the set of the significant climatic variables were observed during the 20th century (Fig. 3 C,D). The effect of summer precipitation has been significant for larch until the mid-part of the 20th century. The effect of temperature in the previous July, August and particularly September has become significant or intensified in moving intervals after about 1950–1990. Precipitation in the current May and temperature in April have been significant for TRW in several intervals, when the trees were younger.
4 Discussion
Good agreement of TRW time series, as shown by EPS > 0.85 and interseries correlation > 0.60 (Table 1), suggested presence of environmental signals in TRW of both species (Wigley et al. 1984). The synchrony between chronologies suggested that both species reacted to similar environmental events; however, a different intensity of reaction was suggested by low correlation between chronologies. The observed signature years (Fig. 2 C) coincided with climate-related pointer years observed for larch in Lithuania (Vitas and Žeimavičius 2010) or beech in Central Europe (Dittmar et al. 2003). Although both species occurred outside their natural range (Bolte et al. 2007; EUFORGEN 2009), a significant but non-drastic effect of the tested climatic variables on TRW was suggested by intermediate correlation/response coefficients (Fig. 2).
Drought related variables generally controlled TRW of both species, as suggested by the negative effect of summer temperature and positive effect of summer precipitation (Fig. 2), as was previously observed in Central Europe (Oleksyn and Fritts 1991; Dittmar et al. 2003). Growth of deciduous trees is partially dependent on nutrient reserves, which are formed at the end of the vegetative period (Barbaroux and Breda 2002; Sudachkova et al. 2004), thus, explaining the effect of climatic conditions in the preceding year (Fig. 2). However, response function coefficients suggested that climatic conditions in the previous year had stronger effect on beech than on larch. Observed changes in TRW-climate relationships and the intensification of the effect of drought-related variables, (Fig. 3) might be explained by warming or alternatively by aging of trees (Carrer and Urbinati 2004). The positive effect of April/May temperature was also observed (Fig. 2), but apparently, it had likely weakened since the 1950s (Fig. 3).
5 Conclusions
Tree-ring width of larch and beech in western Latvia was not narrow (suppressed), suggesting suitability of growth conditions. Climate-TRW analysis suggested that increment was generally limited by climatic variables related to summer drought; a negative effect of temperature and positive effect of precipitation was observed. The expected effect of winter-spring temperature has lost significance, suggesting successful acclimation of the studied trees in stands growing northwards from their natural distribution.
Acknowledgements
The study was funded by Forest Competence Centre (ERAF) project “Methods and technologies for increasing forest capital value” (No. L-KC-11-0004) and Latvian Council of Sciences project “Adaptive capacity of forest trees and possibilities to improve it” (No 454/2012). We also acknowledge Kārlis Taukačš and Juris Kalniņš for help during the sampling and Didzis Elferts, who helped with arrangement of climatic data.
References
Avotniece Z., Rodinov V., Lizuma L., Briede A., Kļaviņš M. (2010). Trends in frequency of extreme climate events in Latvia. Baltica 23: 135–148.
Barbaroux C., Breda N. (2002). Contrasting distribution and seasonal dynamics of carbohydrate reserves in stem wood of adult ring-porous sessile oak and diffuse-porous beech trees. Tree Physiology 22: 1201–1210. http://dx.doi.org/10.1093/treephys/22.17.1201.
Biondi F., Waikul K. (2004). DENDROCLIM2002: a C++ program for statistical calibration of climate signals in tree ring chronologies. Computers and Geosciences 30: 303–311. http://dx.doi.org/10.1016/j.cageo.2003.11.004.
Bolte A., Czajkowski T., Kompa T. (2007). The north-eastern distribution range of European beech – a review. Forestry 80: 413–429. http://dx.doi.org/10.1093/forestry/cpm028.
Bunn A.G. (2008). A dendrochronology program library in R (dplR). Dendrochronologia 26: 115–124. http://dx.doi.org/10.1016/j.dendro.2008.01.002.
Carrer M., Urbinati C. (2004). Age-dependent tree-ring growth response to climate in Larix decidua and Pinus cembra. Ecology 85: 730–740. http://dx.doi.org/10.1890/02-0478.
Cook E.R., Holmes R.L. (1986). Guide for computer program ARSTAN. In: Holmes R.L., Adams R.K., Fritts H.C. (eds.). Tree-ring chronologies of Western North America: California, eastern Oregon and northern Great Basin. University of Arizona, Tucson. p. 50–65.
Dittmar C., Zech W., Elling W. (2003). Growth variation of common beech (Fagus sylvatica L.) under different climatic and environmental conditions in Europe – a dendrochronological study. Forest Ecology and Management 173: 63–78. http://dx.doi.org/10.1016/S0378-1127(01)00816-7.
Dreimanis A. (1995). [The beech and the larch-tree in Shkede forestry]. Dižskābardis un lapegle Šķēdes mežniecībā. In: Forestry education 75th anniversary conference proceedings of Latvia University of Agriculture. p. 94–97.
EUFORGEN (2009). Distribution maps. http://www.euforgen.org/distribution-maps/. [Cited 1 Jan 2014].
Fritts H.C. (2001). Tree-rings and climate. The Blackburn Press, Caldwell. 582 p.
Grissino-Mayer H.D. (2001). Evaluating crossdating accuracy: a manual and tutorial for the computer program COFECHA. Tree-Ring Research 57: 205–221.
Hickler T., Vohland K., Feehan J., Miller P.A., Smith B., Costa L., Giesecke T., Fronzek S., Carter T.R., Cramer W., Kühn I., Sykes M.T. (2012). Projecting the future distribution of European potential natural vegetation zones with a generalized, tree species-based dynamic vegetation model. Global Ecology and Biogeography 21: 50–63. http://dx.doi.org/10.1111/j.1466-8238.2010.00613.x.
Jones P.D., New M., Parker D.E., Martin S., Rigor I.G. (1999). Surface air temperature and its variations over the last 150 years. Reviews of Geophysics 37: 173–199. http://dx.doi.org/10.1029/1999RG900002.
Kullman L. (2008). Thermophilic tree species reinvade subalpine Sweden – early responses to anomalous late Holocene climate warming. Arctic Antarctic and Alpine Research 40: 104–110. http://dx.doi.org/10.1657/1523-0430 (06-120)[KULLMAN]2.0.CO;2.
Lindner M., Maroschek M. et al. (2010). Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. Forest Ecology and Management 259: 698–709. http://dx.doi.org/10.1016/j.foreco.2009.09.023.
Lizuma L., Kļaviņš M., Briede A., Rodinovs V. (2007). Long-term changes of air temperature in Latvia. In: Kļaviņš M. (ed.). Climate change in Latvia. University of Latvia, Riga. p. 11–20.
Oleksyn J., Fritts H.C. (1991). Influence of climatic factors upon tree rings of Larix decidua and L. decidua × L. kaempferi from Pulawy, Poland. Trees-Structure and Function 5: 75–82. http://dx.doi.org/10.1007/BF00227488.
Sudachkova N.E., Milyutina I.L., Romanova L.I., Semenova G.P. (2004). The annual dynamics of reserve compounds and hydrolytic enzymes activity in the tissues of Pinus sylvestris L. and Larix sibirica Ledeb. Eurasian Journal of Forest Research 7: 1–10.
Vetaas O.R. (2002). Realized and potential climate niches: a comparison of four Rhododendron tree species. Journal of Biogeography 24: 545–554. http://dx.doi.org/10.1046/j.1365-2699.2002.00694.x.
Vitas A., Žeimavičius K. (2010). Regional tree-ring chronology of European larch (Larix decidua Mill.) in Lithuania. Baltic Forestry 16: 187–193.
Walther G.R., Post E. et al. (2002). Ecological response to recent climate change. Nature 416: 389–395. http://dx.doi.org/10.1038/416389a.
Wigley T.M.L., Briffa K.R., Jones P.D. (1984). On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. Journal of Climate and Applied Meteorology 23: 201–213. http://dx.doi.org/10.1175/1520-0450(1984)023<0201:OTAVOC> 2.0.CO;2.
Total of 23 references
